
LOCOMOTOR SYNERGY MODIFICATION DURING TREADMILL RUNNING OF VARYING INTENSITY
МОДИФИКАЦИЯ ЛОКОМОТОРНОЙ СИНЕРГИИ ПРИ БЕГЕ РАЗЛИЧНОЙ ИНТЕНСИВНОСТИ ПО ТРЕДБАНУ
JOURNAL: «SCIENTIFIC NOTES OF V.I. VERNADSKY CRIMEAN FEDERAL UNIVERSITY. Biology. Chemistry» Volume 10 (76), №4, 2024
Publication text (PDF):Download
UDK 612.743
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Moiseev S. A., Velikie Luki State Academy of Physical Education and Sports, Velikie Luki, Russia
Mikhailova E. A., Velikie Luki State Academy of Physical Education and Sports, Velikie Luki, Russia
Ershov V. Yu., Velikie Luki State Academy of Physical Education and Sports, Velikie Luki, Russia
TYPE:Article
DOI:https://doi.org/10.29039/2413-1725-2024-10-4-159-167
PAGES:from 159 to 167
STATUS:Published
LANGUAGE:Russian
KEYWORDS: muscle synergies, running, motor control, factor analysis, electromyography.
ABSTRACT (ENGLISH):
The aim of the work was to study the spatial-temporal structure of extensive muscle synergies in conditions of increasing intensity of locomotion. The specific task was to consider changes in the degree of involvement of skeletal muscles in synergy and patterns of their temporary organization in the structure of a full (two-way) cycle of running step with different intensity of work performed.
The study involved 12 athletes specializing in short-distance running. They performed treadmill running in four intensity zones determined by the magnitude of the heart rate – low, moderate, medium, high. Electromyograms (EMG) of 16 superficial bilateral muscles of the upper, lower extremities and trunk were recorded during running. To obtain the parameters of muscle synergies, the method of matrix decomposition by factor analysis using the principal component method without factor rotation was used. The average EMG amplitude, the number of extracted factors, the percentage of total variance taken into account by each factor in the total data set, weighting factors and activation coefficients were analyzed.
It is established that the intermuscular interaction in the structure of the full cycle of a running step can be described by five basic temporal synergies. The main synergies (first and second) demonstrate the stability of the time structure regardless of the intensity of locomotor activity. The main peaks of the patterns of temporary activation of muscle synergies clearly correlate with the periods of manifestation of the greatest electrical activity by the leading muscles. Modification of the spatial organization of muscle synergies is manifested in a significant change in the degree of involvement of individual muscles in the structure of muscle modules with increasing intensity of the work performed.
ВВЕДЕНИЕ
Модульная концепция организации элементов двигательной системы человека предполагает объединение управляемых элементов (мышц) в малоразмерные модули и управление такими образованиями посредством центральных управляющих сигналов [1]. Архитектура таких модулей может охватывать обширные нейрональные сети спинного мозга, поскольку каждая мышца, принимающая участие в локомоторном акте, может иметь представительство в популяциях мотонейронов, локализованных в разных сегментах спинного мозга. Кроме того, отдельный мотонейронный пул может управлять несколькими мышцами. Синергия является механизмом, осуществляющим сопряжение различных двигательных модулей, локализованных на разных уровнях спинного мозга, превращая их в управляемую систему.
При исследовании структуры цикла шага при ходьбе и беге с невысокой скоростью сообщается о структурировании моторного выхода спинальных генераторов в виде пяти базовых компонент, описывающих характер межмышечного взаимодействия в строгом соответствии с периодами шагательного цикла [2]. Однако, несмотря на высокую стереотипность генерируемого локомоторного паттерна на биомеханическом уровне наблюдается довольно большая вариативность пространственной организации паттернов межмышечного взаимодействия. Стереотипность и стабильность временных мышечных паттернов отражает наличие единой стратегии управления локомоцией человека, генерируемой центральными структурами нервной системы, прежде всего на спинальном уровне [3–5]. Однако, каким образом формируется стереотипная временная структура синергий при столь вариативных паттернах пространственной их организации во многом остается не ясным. Выяснение закономерностей формирования и модификации синергий в различных условиях локомоций имеет важное значение для развития общей теории управления произвольными движениями человека и может быть полезным для разработки средств повышения эффективности тренировочного процесса спортсменов, специализирующихся в различных легкоатлетических дисциплинах. Целью работы явилось изучение пространственно-временной структуры обширных мышечных синергий в условиях возрастания интенсивности локомоций. Конкретная задача заключалась в рассмотрении изменений степени вовлечения скелетных мышц в синергии и паттернов их временной организации в структуре полного (двустороннего) цикла бегового шага при разной интенсивности выполняемой работы.
МАТЕРИАЛЫ И МЕТОДЫ
В исследовании приняли участие 12 спортсменов, специализирующихся в беге на короткие дистанции, спортивной квалификации от 1 взрослого разряда до КМС. Возраст испытуемых составлял 18–24 года. Они выполняли бег на тредбане в четырех зонах интенсивности, определяемых величиной частоты сердечных сокращений (ЧСС). Контроль ЧСС осуществляли при помощи нагрудного датчика Polar (Финляндия). Бег, сопровождающийся работой сердца в диапазоне ЧСС 115–125 уд/мин, считали низкой интенсивностью выполняемой работы, 135–145 – умеренной, 155–165 – средней и 175–185 – высокой. Длительность бега в каждой зоне интенсивности определялась с учетом того, чтобы испытуемый совершил не менее 20 полных циклов бегового шага, 12 из которых включали в анализ. В среднем время бега в каждой зоне интенсивности не превышала 40 секунд. Общая длительность бега с учетом времени достижения ЧСС каждой зоны интенсивности и времени выполнения нагрузки не превышала 8 минут.
Во время бега регистрировали электромиограммы (ЭМГ) 16 поверхностных билатеральных мышц верхних, нижних конечностей и туловища: дельтовидной (DALTA), выпрямляющей позвоночник (ES), прямой живота (RA), большой ягодичной (GM), прямой мышцы бедра (RF), двуглавой мышцы бедра (BF), икроножной (MG), передней большеберцовой (TA). Запись ЭМГ осуществлялась биомонитором МЕ 6000 (Финляндия) с частотой оцифровки 2000 Гц. ЭМГ фильтровали полосовым фильтром 30–450 Гц, усредняли в интервалах 0,002 с. Синхронно с ЭМГ посредством системы «Qualisys» (Швеция) осуществляли видеозахват основных антропометрических точек сегментов тела. Период опоры определялся от постановки конечной антропометрической точки правой ноги на поверхность ленты тредбана и до ее отрыва, период переноса определялся от отрыва данной точки до постановки ее на ленту тредбана.
Зарегистрированные данные ЭМГ и координаты антропометрических точек экспортировали в Statistica (StatSoft, Inc., version 10), где формировали матрицу, состоящую из вариационных рядов регистрируемых величин и дополнительных переменных, идентифицирующих принадлежность измерений к различным испытуемым, беговым циклам, периодам бегового шага и пр. [6]. Разложение матриц осуществляли с помощью факторного анализа по методу главных компонент без вращения факторов. Исходная матрица X разлагалась на произведение двух матриц: X=T×P+E, где T – матрица счетов, P – матрица нагрузок, E – матрица остатков.
Матрица нагрузок (Р) несет информацию о взаимосвязи или независимости переменных относительно новых, формальных переменных, полученных в процессе разложения матриц – «весовые коэффициенты», чем выше коэффициент, тем больше связь с новой синергией. Весовой коэффициент более 0,70 считали высоким, от 0,35 до 0,70 – средним, менее 0,35 – низким. Матрица счетов (Т) определяет временную организацию выявленных синергий и представляет собой проекции исходных данных на подпространство главных синергий – «коэффициенты активации», т.е. процесс, показывающий изменение мышечной активности каждого из факторов во времени. Использовали стандартный модуль Statistica «Mult/Exploratory – Factor». Критериями определения количества выделяемых факторов служили показатели собственных значений (>1) и процент учитываемой дисперсии каждым компонентом (не менее 5 %).
Анализировали среднюю амплитуду ЭМГ, количество извлекаемых факторов, процент общей дисперсии, учитываемый каждым фактором в общем наборе данных, весовые коэффициенты и коэффициенты активации. Статистическая обработка включала расчет среднего арифметического (M), ошибки среднего арифметического (m), стандартного отклонения (SD). Нормальность распределения данных определяли с помощью теста Шапиро-Уилка при P>0,05.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
При беге по тредбану динамика показателей ЭМГ скелетных мышц носила волнообразный характер, периоды относительно низкой электроактивности сменялись кратковременными периодами повышения амплитуды биопотенциалов (рис. 1).
В периоде опоры наибольшую амплитуду электроактивности относительно других мышц демонстрировали медиальная головка икроножной мышцы – 150,18±60,36 мкВ и передняя большеберцовая мышца – 111,29±27,05 мкВ правой ноги, а также двуглавая мышца бедра левой ноги – 138,52±35,20 мкВ. В период переноса ноги наибольшие значения средней амплитуды ЭМГ были получены для левой ягодичной мышцы – 76,37±36,07 мкВ, правой и левой дельтовидных мышц и левой икроножной мышцы (70,13±20,20 мкВ и 96,57±35,71 мкВ). Эти же мышцы проявляли бόльшую электроактивность при изменении интенсивности бега.
Количество извлекаемых синергий при разной интенсивности бега существенно не различалось (P>0,05). При низкой и умеренной интенсивности выполняемой работы их количество достигало шести, а в среднем составляло 5,00±0,40 и 5,25±0,25 соответственно. При средней и высокой интенсивности количество синергий варьировало от четырех до пяти. Процент объясняемой дисперсии компонентами во всех условиях превышал 80 %, что свидетельствует о высоком качестве реконструкции исходных данных.

Рис. 1. Усредненная ЭМГ-активность скелетных мышц в различных периодах бегового шага при разной интенсивности бега. По оси абсцисс – прогресс цикла бегового шага (0 % – начало цикла, 100 % – окончание цикла).
Установлено, что временная структура первой синергии характеризовалась двумя выраженными пиками активации в структуре цикла бегового шага при всех рассматриваемых условиях бега (рис. 2). При разной интенсивности бега первый пик всегда приходился на начало периода опоры, а второй на начало периода переноса. Паттерны активации данной синергии демонстрировали высокое сходство при разной интенсивности локомоций (r>0,8). Структура второй синергии при низкой интенсивности бега имела профиль с двумя пиками активации мышц, приходящихся на те же периоды цикла бегового шага, что и в первой синергии. Однако при увеличении интенсивности бега сохранялся только второй пик в начале периода переноса. Профили активации двух выделенных синергий демонстрировали низкую межиндивидуальную вариативность.
Профили синергий 3–5 оказались более вариативны, однако в их структуре можно выделить основные пики. Чаще в структуре названных синергий выделялось по одному основному пику активации. Например, для третьей синергии пик активации приходился на вторую четверть цикла бегового шага при низкой интенсивности выполняемой работы (см. рис. 2). При увеличении интенсивности локомоций регистрировался один выраженный пик активации, но было отмечено его смещение на временной шкале относительно середины цикла шага в левую или правую сторону.
При изменении интенсивности бега расположение участков, отражающих возрастание электроактивности мышц, на временной шкале оставалось постоянным. Временной период основных пиков активности может быть соотнесен с событиями, характеризующими биомеханическую структуру локомоторного цикла. Так, пик активации первой синергии в первой четверти цикла бегового шага наблюдается при постановке правой ноги на опору и сопровождается возрастанием электроактивности икроножной мышцы. Постановка левой ноги на опору сопровождается возрастанием электроактивности мышц бедра и голени и характерным пиком активации временной синергии (см. рис. 2). Структура основных временных синергий (первая и вторая) и их соотношение с периодами проявления максимальных мышечных усилий сохраняется при изменении интенсивности бега.
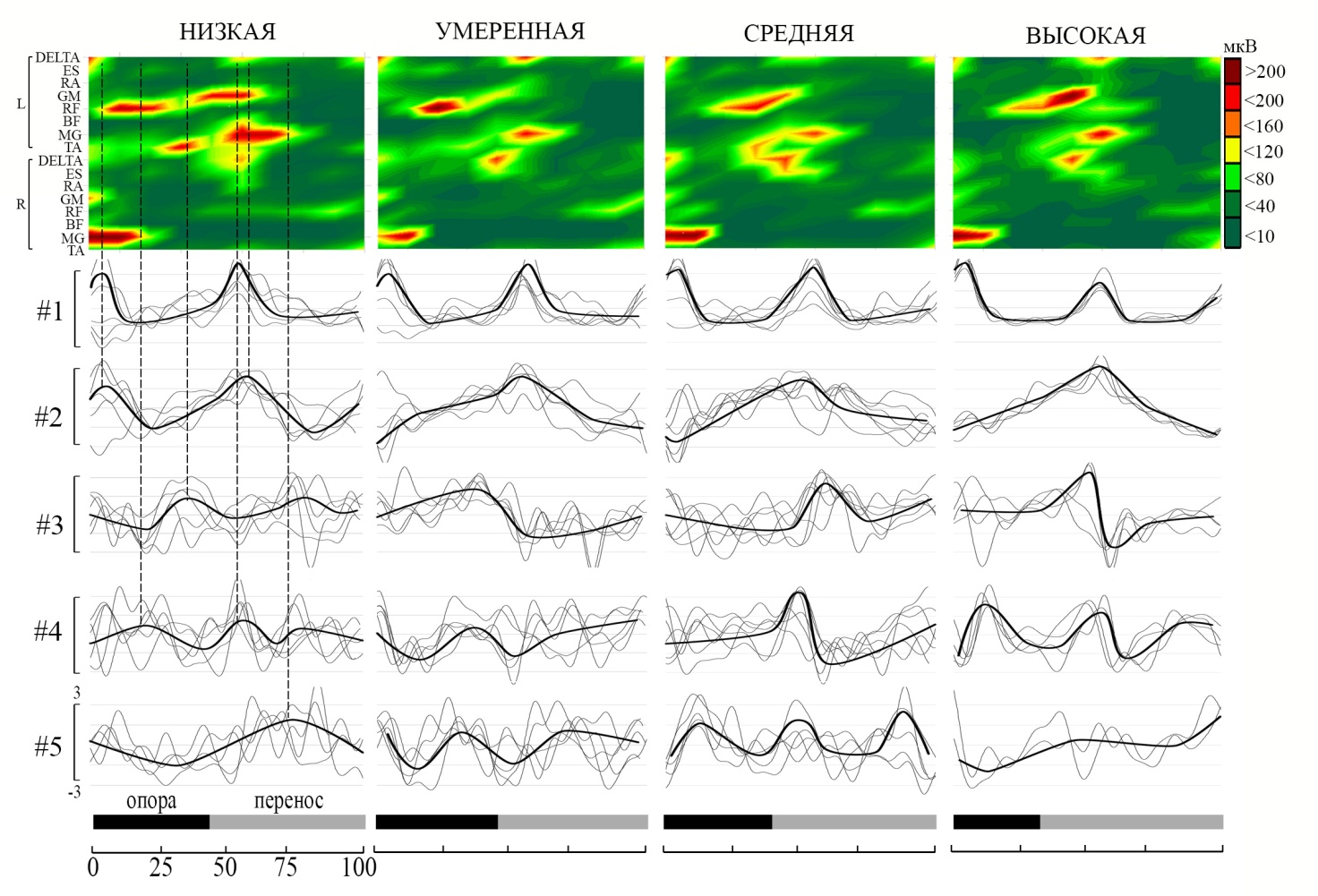
Рис. 2. Динамика средней амплитуды ЭМГ и основные временные синергии мышечной активности при беге различной интенсивности. По оси абсцисс – прогресс цикла бегового шага (0 % – начало цикла, 100 % – окончание цикла). Тонкие линии – усредненные внутрииндивидуальные профили, жирные линии – среднегрупповые профили. #1–5 – синергии.
Многие исследователи указывают на стабильность временной структуры мышечной активации, например, при педалировании на велоэргометре, при ходьбе по тредбану или при обычной ходьбе с разной скоростью даже при использовании разных методов факторизации матриц [3, 7, 8]. Однако, наблюдаются сдвиги основных пиков временной активации синергий при изменении условий реализации двигательной задачи, например, при ходьбе по неровной поверхности или при педалировании с разным уровнем сопротивления [9–13].
Характерной чертой синергии, как способа организации элементов управляемой системы является степень синхронизации мышечных активаций. Такую информацию могут дать, получаемые в результате разложения матриц, весовые коэффициенты каждой мышцы в структуре выделяемых синергий (рис. 3).
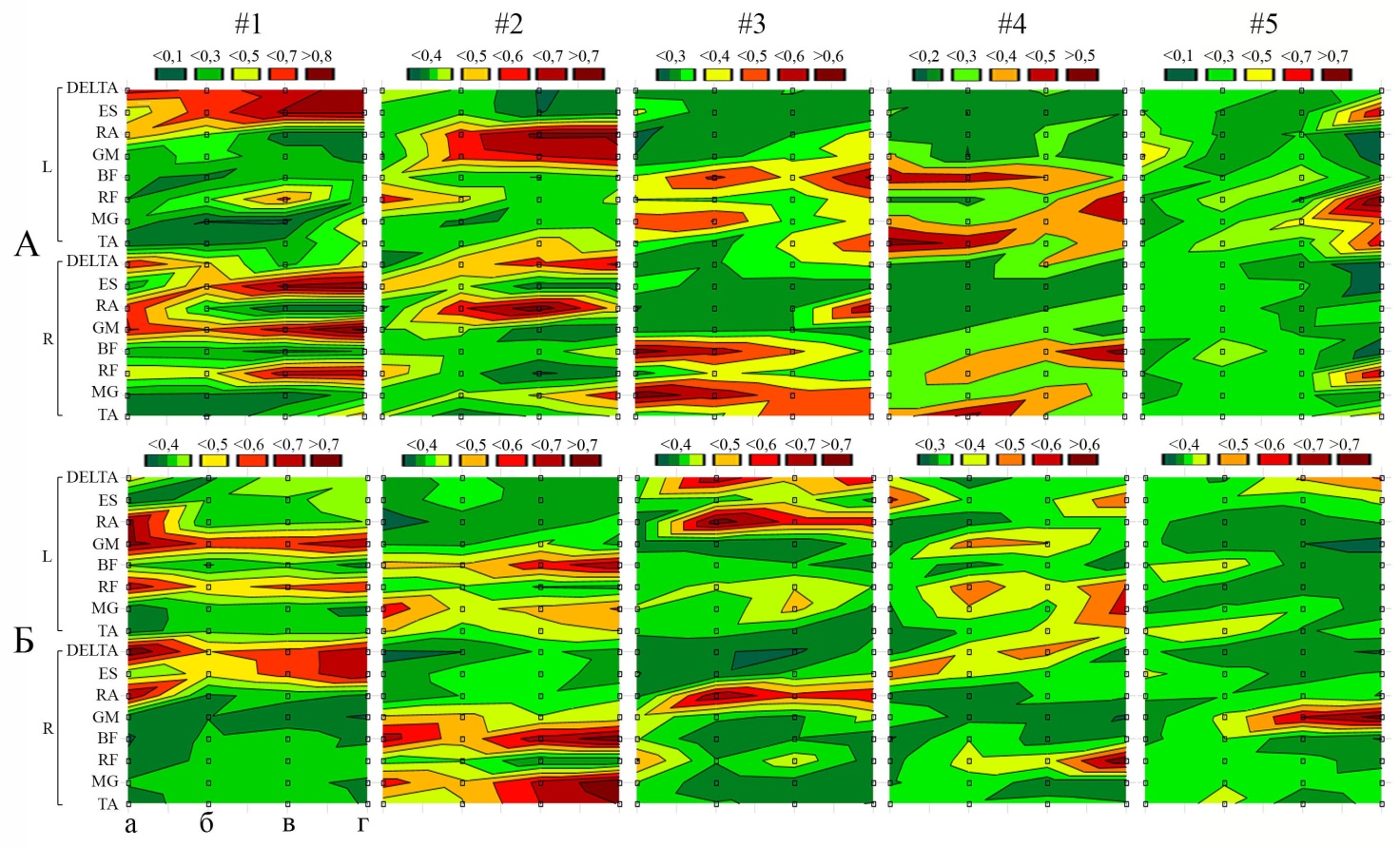
Рис. 3. Весовые коэффициенты мышц в структуре выделяемых синергий. Интенсивность бега: а – низкая, б – умеренная, в – средняя, г – высокая. L – левая сторона, R – правая сторона. #1–5 – синергии. А – период опоры, Б – период переноса ноги.
Первая синергия при низкой интенсивности бега в периоде опоры включала наибольшие коэффициенты для дельтовидной (0,68±0,07) прямой мышцы живота (0,68±0,07), а также большой ягодичной мышцы правой стороны тела – 0,71±0,04. В периоде переноса наибольших значений достигали коэффициенты левой прямой мышцы бедра, правой дельтовидной мышцы, прямой мышцы живота обеих сторон тела, здесь значения находились в диапазоне от 0,65±0,03 до 0,78±0,04. Вторая синергия при той же интенсивности бега характеризовалась средними и низкими весовыми коэффициентами мышц, а наибольших значений достигали прямая мышца бедра (0,54±0,07) и мышца, выпрямляющая позвоночник (0,48±0,05) правой стороны тела. При переносе ноги наибольшие весовые коэффициенты были получены для двуглавой мышцы бедра – 0,64±0,06 и икроножных мышц обеих нижних конечностей (0,54±0,03 и 0,56±0,04). Синергии 3–5, преимущественно, включали одну и реже две мышцы, демонстрирующие высокие коэффициенты в обоих рассматриваемых периодах цикла бегового шага.
ЗАКЛЮЧЕНИЕ
Межмышечное взаимодействие в структуре полного цикла бегового шага может быть описано пятью базовыми временными синергиями. Основные синергии (первая и вторая) демонстрируют стабильность временной структуры вне зависимости от интенсивности локомоторной активности. Основные пики паттернов временнόй активации мышечных синергий четко соотносятся с периодами проявления наибольшей электроактивности ведущими мышцами. Модификация пространственной организации мышечных синергий проявляется в существенном изменении степени вовлечения отдельных мышц в структуру мышечных модулей при возрастании интенсивности выполняемой работы.
- D’Avella A. Modularity for Motor Control and Motor Learning. Advances in Experimental Medicine and Biology, 957, 3 (2016)
- Ivanenko Y., Poppele R., Lacquaniti F. Five basic muscle activation patterns account for muscle activity during human locomotion. J Physiol, 556 (1), 267 (2004).
- Hug F., Turpin N. A., Guével A., Dorel S. Is interindividual variability of EMG patterns in trained cyclists related to different muscle synergies? Journal of Applied Physiology, 108 (6), 1727 (2010).
- De Marchis C., Schmid M., Bibbo D., Bernabucci I., Conforto S. Inter-individual variability of forces and modular muscle coordination in cycling: a study on untrained subjects. Hum Mov Sci, 32 (6), 1480 (2013).
- Esmaeili J., Maleki A. Comparison of muscle synergies extracted from both legs during cycling at different mechanical conditions. Australas Phys Eng Sci Med, 42 (3), 827 (2019).
- Moiseev S. A., Pukhov A. M., Mikhailova E. A., Gorodnichev R. M. Methodological and Computational Aspects of Extracting Extensive Muscle Synergies in Moderate-Intensity Locomotions. Journal of Evolutionary Biochemistry and Physiology, 58 (1), 88 (2022).
- Saito A., Tomita A., Ando R., Watanabe K., Akima H. Similarity of muscle synergies extracted from the lower limb including the deep muscles between level and uphill treadmill walking. Gait Posture, 59, 134 (2018).
- Rabbi M. F., Pizzolato C., Lloyd D. G., Carty C. P., Devaprakash D., Diamond L. E. Non-negative matrix factorization is the most appropriate method for extraction of muscle synergies in walking and running. Sci Rep., 10 (1), 8266 (2020).
- Moiseev S. A., Gorodnichev R. M. Motor Synergies Structure Variability in Different Intensity Locomotions. Fiziologiya cheloveka, 48 (4), 22 (2022) (in Russ).
- Mileti I., Serra A., Wolf N., Munoz-Martel V., Ekizos A., Palermo E., Arampatzis A., Santuz A. Muscle Activation Patterns Are More Constrained and Regular in Treadmill Than in Overground Human Locomotion. Front Bioeng Biotechnol., 8, 619 (2020).
- Santuz A., Brüll L., Ekizos A., Schroll A., Eckardt N., Kibele A., Schwenk M., Arampatzis A. Neuromotor Dynamics of Human Locomotion in Challenging Settings. iScience., 23 (1), 1 (2020).
- Santuz A., Ekizos A., Kunimasa Y., Kijima K., Ishikawa M., Arampatzis A. Lower complexity of motor primitives ensures robust control of high-speed human locomotion. Heliyon., 6 (10), 1. (2020).
- Nishida K., Hagio S., Kibushi B., Moritani T., Kouzaki M. Comparison of muscle synergies for running between different foot strike patterns. PLoS One, 12 (2), 1 (2017).
