
MORPHOFUNCTIONAL INDICATORS OF MILITARY PERSONNEL WITH AMPUTATION OF LOWER LEG AT THE STAGE OF PREPARATION FOR PROSTHETICS
ОЦЕНКА МОРФОФУНКЦИОНАЛЬНОГО СОСТОЯНИЯ ВОЕННОСЛУЖАЩИХ С АМПУТАЦИЕЙ ГОЛЕНИ НА ЭТАПАХ ФИЗИЧЕСКОЙ РЕАБИЛИТАЦИИ
JOURNAL: «SCIENTIFIC NOTES OF V.I. VERNADSKY CRIMEAN FEDERAL UNIVERSITY. Biology. Chemistry» Volume 10 (76), №2, 2024
Publication text (PDF):Download
UDK: 575.21 (634.2: 581.45, 581.47)
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Anatov D. M., Precaspian Institute of Biological Resources of the Dagestan Federal Research Centre of the RAS, Makhachkala, Russia
Gabibova M. R., Precaspian Institute of Biological Resources of the Dagestan Federal Research Centre of the RAS, Makhachkala, Russia
TYPE:Article
DOI:https://doi.org/10.29039/2413-1725-2024-10-2-3-13
PAGES:from 3 to 13
STATUS:Published
LANGUAGE:Russian
KEYWORDS: Microcerasus incana, variability of morphological traits, structure of correlations, fruits, leaves, Dagestan.
ABSTRACT (ENGLISH):
Based on the results of the study, new data were obtained on phenotypic variability and the structure of correlations of morphological characteristics of the fruit and leaves of M. incana in the conditions of Dagestan. For the first time, the results of the interaction of coordinated and general variability of morphological traits of the fruit and leaf are presented. The range of intrapopulation variation in fruit and leaf traits has been established.
Analysis of intrapopulation variability showed that the average fruit diameter is 7.4 mm with a range of 5.9–9.9 mm, and the endocarp size is 4.7 mm. The percentage of stones in the fruit averaged 18.8 %. The length of the leaf is on average 43.5 mm, varying from 28 to 61 mm, while the width of the leaf is three times less than the length, the leaf index was 31.0 % and fluctuates in the range.
A wide range of variability in fruit and leaf traits was identified based on the coefficient of variation (CV). All geometric traits of the fruit and endocarp, leaf length, and petiole diameter were characterized by low variation. The weight traits of the fruit, petiole length, weight and leaf area were characterized by high variations. Index traits for highly correlated characters showed lower variability compared to the morphological traits that made up the index, these include SLA, fruit shape and endocarp indices, endocarp yield, and high for weakly correlated ones.
Correlation analysis showed that out of 210 possible variants of correlation relationships of the considered features, 51 turned out to be reliable. Visual clustering of the correlation structure using the Früchterman-Reingold algorithm revealed two clearly separate correlation graphs corresponding to their structures. Low reliable positive relationships were established between the characteristics of the leaf and the fruit (r = 0.36–0.39); they were noted for the pairs “fruit mass” – “petiole diameter”, “fruit mass” – “leaf mass” and “petiole diameter” – “length” bones.” Functional connections between linear characteristics of the fruit and endocarp, mass and leaf area, and others were identified. No negative significant relationships were identified. Strong correlations were noted between leaf length and perimeter (r=0.94), leaf width and area (r=0.88). The main connecting characteristics between the structures are the diameter of the petiole and the weight of the fruit.
Analysis of the structure of variability by (CV) and determination (R2ch) showed that group I (ecological-biological system indicators) consisted of the traits fruit and endocarp weight, petiole length and diameter. Group II (biological indicators) included all linear signs of the fruit and seeds. Group III (genotypic indicators) was formed by the length, width and leaf weight. Group IV (ecological system indicators) includes leaf area and perimeter.
ВВЕДЕНИЕ
Дикорастущие виды плодовых культур издавна привлекали внимание ботаников и практиков растениеводов [1]. В свое время выдающийся генетик и селекционер Н. И. Вавилов придавал огромное значение изучению и использованию огромного многообразия диких сородичей плодовых культур в селекции [2]. На Кавказе и в Дагестане наибольшее разнообразие видов относится к подсемейству Prunoideae семейства Rosaceae. Это такие виды, как например: Prunus armeniaca L. – Абрикос обыкновенный, P. cerasifera Ehrh. – Алыча растопыренная, P. spinosa L. – Терн, Cerasus avium (L.) Moench – черешня, Microcerasus incana (Pall.) M. Roem. – Микровишня седая и др.
Как и большинство видов рода Prunus этот вид вовлечен в селекционный процесс, где особое значение отводится созданию новых клоновых подвоев для косточковых благодаря высокой зимостойкости и засухоустойчивости из всех вишен. Кроме этого, этот вид нетребователен к почве, мало повреждается вредителями и болезнями. Может представлять интерес для декоративного озеленения за счет раннего обильного цветения и красочному плодоношению [3].
Для генетико-ресурсоведческих целей изучение внутривидовой изменчивости имеет большое значение если проводится анализ причин изменчивости [4]. Одним из основных методов оценки изменчивости является анализ взаимосвязи физиологических, анатомических и морфологических признаков исследуемого объекта с их показателями общей и согласованной изменчивости [5, 6].
В этой связи изучение структуры изменчивости морфологических признаков в естественных условиях вишни седой (Microcerasus incana (Pall.) Roem.) может способствовать решению ряда вопросов микроэволюции, экологии и ресурсоведения. Все это создает предпосылки для всестороннего изучения вишни седой как перспективного ресурсного вида в селекционных программах.
Цель работы – определить характер взаимосвязи согласованной и общей изменчивости морфологических признаков плода и листа M. incana на примере отдельно взятой популяции в условиях Дагестана.
МАТЕРИАЛЫ И МЕТОДЫ
Microcerasus incana (Pall.) Roem (Микровишня седая) – невысокий листопадный кустарник высотой 1–2 м, с плотной раскидистой кроной. Кора старых побегов светло-коричневая с желтыми чечевичками, у молодых побегов – серая, переходящая к верхушке в красно-коричневую (рис. 1). Листья длиной до 5 см продолговатые или продолговато ланцетные или узко-обратнояйцевидные, сверху голые или коротковолосистые, снизу беловойлочные на небольших, короткоопушенных черешках. Край листовой пластинки широко-острозубчатый. Прилистники шиловидные, большей частью перистонадрезанные. Цветки почти сидячие до 1,5 см в диаметре, розово-красные. Доли чашечки значительно короче цветоложа. Продолжительность цветения 5–16 дней. Костянка маленькая, темно-красная, яйцевидно-шаровидная. Основной ареал вида – Северный Кавказ, Закавказье, Малая Азия, Средняя Азия, Западная Сибирь, и на Балканах. Светолюбивый ксерофит, мезотерм, мезотроф [7, 8]. В Дагестане этот вид широко распространён в горной части от 300 до 1500 (1700) н. у. м., встречаясь разреженно небольшими локальными группами. Предпочитает открытые сухие склоны и заросли кустарников.
В качестве учетной модельной популяции для оценки фенотипического разнообразия и внутрипопуляционной изменчивости морфологических признаков плода и листа была выбрана популяция в условиях Внутригорного Дагестана, произрастающая в Губденском ущелье (Карабудахкентский район, долина р. Пулеувла, h=440 м н. у. м.). Данная популяция произрастает на щебнистых известняковых породах склонах южных экспозиций. Почвенный покров не выражен. В отмеченной популяции средняя годовая температура – +8,2 – +10,5 °С, температура летнего периода – +15,5 – +22 °С, зимнего периода – –0 – –4 °С. Характерной особенностью типичных мест произрастания вишни седой является невысокое годовое количество осадков (460–550 мм за год), в основном, выпадающих в период активной вегетации (май-июль) [9].

Рис. 1. Кустарник Microcerasus incana Губденской популяции.
Камеральная обработка проведена у 30-ти случайно отобранных кустарников, находящихся в зрелой генеративной стадии. При описании использовалось десять типичных плодов и листьев. Были учтены 8-ми континуальных размерных и весовых, 7-ми индексных признаков плода и эндокарпия (косточки). Сравнение особей проведена по следующим количественным характеристикам: плоды – длина (FL), ширина (FW), толщина (FT), общая масса (FM), эндокарпий – длина (КL), ширина (КW), толщина (КT), масса (КМ), индексы плода: выход косточки в процентах (Ra=КM/FM), геометрический средний диаметр плода (DgF =√(FL+FW+FT)/3) и эндокарпия (DgS =√(КL+КW+КT)/3).Из количественных признаков измерялись длина (LL) и ширина (LW) листовой пластинки, длина (PL) и диаметр (PD) черешка, масса (LM), площадь (P) и периметр (PL) листовой пластинки. Дополнительно были рассчитаны три соотносительных показателя:
If – листовой индекс (LW*100%/LL), SLA – удельная площадь листа (P/LM),
Ip – черешковый индекс (LL/PL).
Для каждого признака находились: среднее арифметическое (Х), ошибка среднего (Sх), стандартное отклонение (S). Относительную изменчивость изучаемых признаков проводили по значению коэффициента вариации (CV, %) с учётом шкалы изменчивости [10]. Оценку структуры изменчивости морфологических признаков производилась с помощью сопоставления коэффициента вариации (СV, %) и коэффициента детерминации (квадрат коэффициента корреляции, усредненный по всем признакам – R2ch) [6].
Для анализа корреляционной структуры морфологических признаков (без индексов) была применена визуальная их кластеризация с использованием алгоритма Фрюхтермана-Рейнгольда (Fruchterman-Reingold).
Статистический анализ результатов измерений проведен с использованием программ Statistica v.13.3. График корреляционной структуры осуществлен в программе RStudio [11].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Анализ количественных признаков плода вишни седой в условиях Губденского ущелья показало, что размеры колеблются в широких пределах. Размах варьирования по длине плода составила 5,7–9,2 мм, по ширине 5,6–9,8 мм и 6–10,8 мм по толщине, что можно охарактеризовать учтенную популяцию как гетерогенную (табл. 1). Более обобщенную картину размерным признакам плода придает индексный показатель DgF – геометрический диаметр плода, измеренный по всем направлениям, который варьирует в пределах 5,9–9,9 мм и в среднем составляет 7,4 мм. Размеры косточек в диапазоне от 4,1 до 7,1 мм, в среднем 5,6 мм. Ширина и толщина эндокарпия значительно уступают длине и в среднем составили 4,5 и 4,1 мм соответственно. Косточка в плодах по индексу Ra может занимать от 12,1 до 33,3 % и в среднем 18,8 %. Средняя масса плода составляет 329,8 мг, а косточки 61,0 мг.
Амплитуда изменчивости по коэффициенту вариации (CV) характеризуются низким варьированием для линейных признаков плода и эндокарпия и высоким для весовых.
Таблица 1
Суммарные статистики признаков плода и эндокарпия M. incana
|
Признаки (n=300) |
Х±Sx |
Min–max |
CV, % |
|
Длина плода, мм |
7,1±0,04 |
5,7–9,2 |
9,2 |
|
Ширина плода, мм |
7,2±0,04 |
5,6–9,8 |
9,9 |
|
Толщина плода, мм |
8,0±0,05 |
6,0–10,8 |
10,4 |
|
Масса плода, мг |
329,8±5,21 |
164,0–685,0 |
27,3 |
|
Длина эндокарпия, мм |
5,6±0,03 |
4,1–7,1 |
9,4 |
|
Ширина эндокарпия, мм |
4,5±0,02 |
3,6–5,8 |
9,5 |
|
Толщина эндокарпия, мм |
4,1±0,02 |
3,0–5,2 |
8,5 |
|
Масса эндокарпия, мг |
61,0±0,87 |
29,0–114,0 |
24,7 |
|
Ra, % |
18,8±0,18 |
12,1–33,3 |
16,4 |
|
DgF, мм |
7,4±0,04 |
5,9–9,9 |
9,0 |
|
DgS, мм |
4,7±0,02 |
3,8–5,8 |
8,1 |
Морфологические признаки листа также могут варьировать в широких пределах значений (табл. 2). Длина листовой пластинки варьирует от 28 до 61 мм и в среднем составляет 43,5 мм. Ширина листа примерно в 3 раза меньше длины и колеблется в диапазоне 6,0–21 мм.
Показатель SLA в среднем составило 70,0 см2/г, что в целом характеризует этот вид как светолюбивый [12–14]. Относительно низкие значения его у вида возможно связано с произрастанием в аридных условиях. Также более мелкие размеры листьев могут быть характерны для видов из более холодных и бедных питательными веществами местообитаний [15].
По коэффициенту вариации наиболее стабильными признаками оказались длина пластинки и диаметр черешка. Сильным варьированием характеризовались длина черешка, масса и площадь листа.
Таблица 2
Суммарные статистики признаков листа M. incana
|
Признаки (n=300) |
Х±Sx |
Min–max |
CV, % |
|
Длина пластинки, мм |
43,5±0,36 |
28,0–61,0 |
14,4 |
|
Длина черешка, мм |
2,7±0,05 |
1,0–5,0 |
32,5 |
|
Диаметр черешка, мм |
0,7±0,01 |
0,4–1,0 |
15,0 |
|
Ширина листа, мм |
13,5±0,17 |
6,0–21,0 |
22,1 |
|
Периметр листа, см |
12,3±0,11 |
7,6–16,7 |
15,4 |
|
Площадь листа, см2 |
4,6±0,08 |
1,6–8,2 |
29,0 |
|
Масса листа, мг |
66,2±1,10 |
20,0–117,0 |
28,8 |
|
Листовой индекс, % |
31,0±0,34 |
20,4–56,3 |
19,1 |
|
SLA, см2/г |
70,0±0,47 |
50,5–94,0 |
11,7 |
|
Черешковый индекс |
4,0±0,07 |
1,2–8,2 |
32,3 |
Линейные размеры разных структур растений, могут варьировать в широких пределах на внутри- и межиндивидуальном уровне по различным причинам, но их относительные размеры (индексы), могут характеризоваться большим постоянством при условии, что составляющие индекс признаки высокоскоррелированы [16].
Анализ изменчивости количественных признаков плода и листа по коэффициенту вариации (CV) на общепопуляционном уровне показал, что почти всех индексные признаки показали более низкую изменчивость по сравнению с морфологическими признаками составлявшие индекс, это: SLA, геометрические размеры плода и эндокарпия, и выход косточки (табл. 1, 2). Индексы формы листа и черешка характеризовались промежуточной изменчивостью, характерной для слабо скоррелированных признаков. Коэффициент вариации отобразил низкое варьирование для индексных признаков плода и эндокарпия, удельной листовой поверхности; среднее для листового индекса; высокое у черешкового индекса.
Проведенный корреляционный анализ для усредненных значений показал, что из 210 возможных вариантов корреляционных взаимосвязей морфологических (без учета индексов) признаков достоверными выделились 51. При сопоставлении отмеченных признаков листа и плода заметно выявляются две корреляционные плеяды, соответствующие их морфоструктурам (рис. 2). Слабые достоверные положительные связи между признаками листа и плода отмечены между «массой плода – FM», «диаметром черешка – PD» и «массой листа – LM». Диаметр черешка также связан с «длиной эндокарпия – KL». Некоторые связи чисто функциональные, например, линейные признаки плода и эндокарпия, масса листа и площадь и др. Таких связей с показателями r ≥ 0,7 оказалось 37 из 51. Отрицательных достоверных связей не было выявлено. Сильная корреляция отмечена между длиной и периметром листа (r=0,94), шириной и площадью листа. Связующими признаками между плеядами можно отнести диаметр черешка и масса плода.
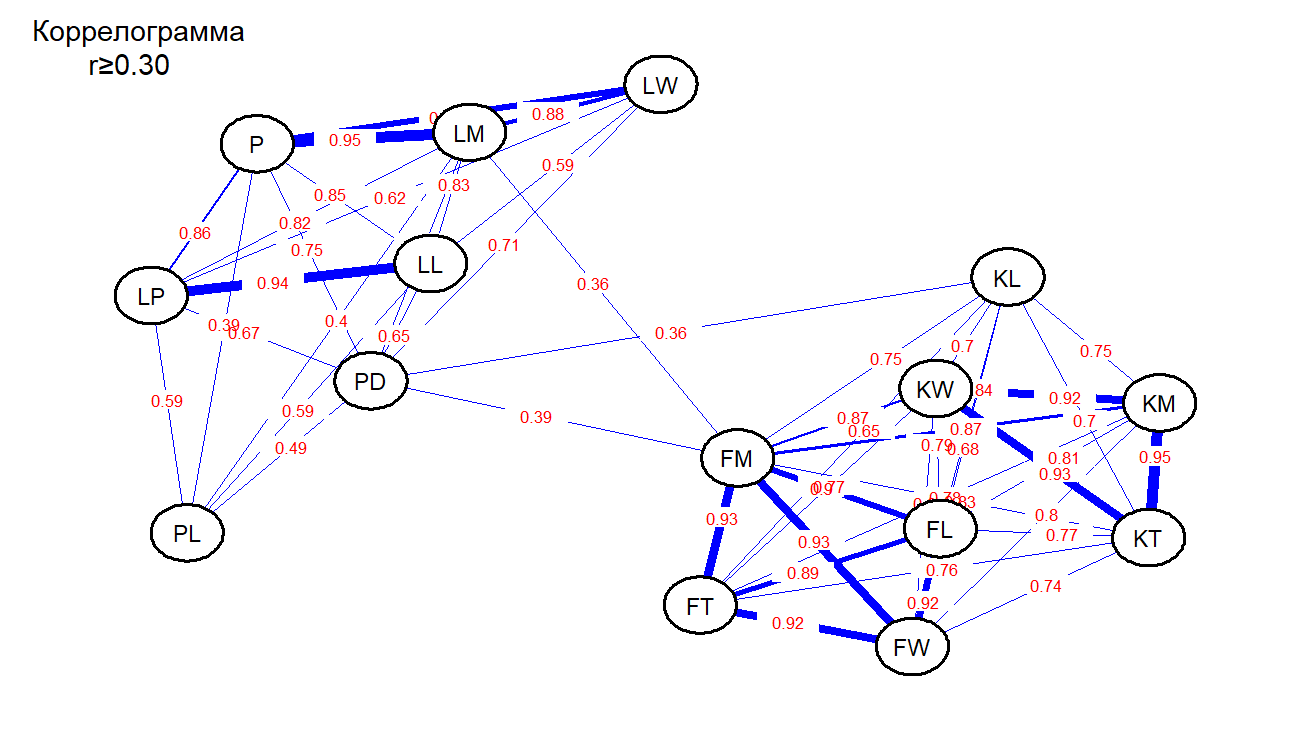
Рис. 2. Структура корреляций признаков плода и листа M. incana.
Результаты исследований структуры изменчивости морфологических признаков по (CV) и детерминации (R2ch) отражающих вклад генотипических факторов и окружающей среды представлено на рис. 3. Анализ структуры изменчивости показал, что в группу I (эколого-биологические системные индикаторы) с относительно высокой детерминированностью и изменчивостью вошли признаки масса плода и эндокарпия, а также длина и диаметр черешка (рис. 3, I). В группу II биологических индикаторов (слабо изменчивые и сильно интегрированные) компактно расположились все линейные признаки плода и эндокарпия (рис. 3, II). В группу III (генотипические индикаторы с наименьшей изменчивостью по CV и R2ch) вошли признаки: длина, ширина и масса листа (рис. 3, III). К группе IV (экологические системные индикаторы) можно отнести площадь и периметр листа (рис. 3, IV). Эти признаки характеризуются высокой относительной изменчивостью и низкой детерминированностью. Полученные данные относительной изменчивости и детерминированности признаков показывают, что самой сильной генотипической автономностью характеризуется длина листа, а типичным индикатором согласованной изменчивости – масса плода. Классификация признаков по группам хорошо согласуется с дифференциаций по морфоструктурам (лист, плод), за исключением группы I, в которую вошли признаки обоих групп.
Рис. 3. Ординация структуры изменчивости морфологических признаков плода и листа M. incana по группам.
ЗАКЛЮЧЕНИЕ
По результатам проведенного исследования получены новые данные по фенотипической изменчивости и структуре корреляций морфологических признаков плода и листа M. incana в условиях Дагестана. Впервые представлены результаты взаимодействия согласованной и общей изменчивости морфологических признаков плода и листа M. incana. Установлен размах внутрипопуляционного варьирования признаков плода и листа.
Анализ внутрипопуляционной изменчивости показал, что средний диаметр плода составляет 7,4 мм с диапазоном 5,9–9,9 мм, эндокарпия – 4,7 мм. Процентное содержание косточки в плоде составила в среднем 18,8 %. Длина листовой пластинки в среднем составляет 43,5 мм, варьируя от 28 до 61 мм, при этом ширина листа в 3 раза меньше длины, листовой индекс составил 31,0 %.
Выявлен широкий спектр изменчивости признаков плода и листа по коэффициенту вариации (CV). Низким варьированием характеризовались все линейные признаки плода и эндокарпия, длина листа, диаметр черешка. Высоким варьированием характеризовались весовые признаки плода, длина черешка, масса и площадь листа. Индексные показатели для функционально скоррелированных признаков выявили более низкую изменчивость по сравнению с морфологическими признаками составлявшие индекс к ним относятся SLA, индексы формы плода и эндокарпия, выход косточки и высокую для слабо скоррелированных.
Корреляционный анализ показал, что из 210 возможных вариантов корреляционных взаимосвязей учтенных признаков достоверными оказались 51. Визуальная кластеризация структуры корреляций с использованием алгоритма Фрюхтермана-Рейнгольда выявила две четко обособленные корреляционные плеяды, группирующиеся внутри морфоструктур (плод, лист). Между признаками листа и плода установлены слабые достоверные положительные связи (r= 0,36-0,39) отмечены для пар «масса плода» – «диаметр черешка», масса плода» – «масса листа» и «диаметр черешка» – «длина эндокарпия». Выделены функциональные связи между линейными признаками плода и эндокарпия, массой и площадью листа и другие. Отрицательных достоверных связей не было выявлено. Сильные корреляции отмечены между длиной и периметром листа (r=0,94), шириной и площадью листа r=0,88). Основными связующими признаками между структурами являются диаметр черешка и масса плода.
Анализ структуры изменчивости по (CV) и детерминации (R2ch) показал, что группу I (эколого-биологические системные индикаторы) составили признаки масса плода и эндокарпия, длина и диаметр черешка; в группу II (биологические индикаторы) вошли все линейные признаки плода и эндокарпия; III группу (генотипические индикаторы) образовали признаки длина, ширина и масса листа; к группе IV (экологические системные индикаторы) можно отнести площадь и периметр листа.
- Yushev A. A., Orlova S. Y. Wild cherries of the Caucasus, Central Asia and the Far East and their use in breeding. Proceedings on Applied Botany, Genetics and Breeding, 180(3), 59 (2019).
- Vavilov N. I. Wild relatives of fruit trees in the Asian part of the USSR and the Caucasus and the problem of the origin of fruit trees. Proceedings on Applied Botany, Genetics and Breeding, 26(3), 85 (1931).
- Eremin G.V., Eremin V.G. Use of the gene pool of wild species of the genus Prunus L. in the selection of clonal rootstocks of stone fruit crops. Proceedings on Applied Botany, Genetics and Breeding, 176(4), 416 (2015).
- Magomedmirzaev M. M. Ways to identify and use genetic resources of higher plants. General Genetics, 3, 130 (1978).
- Rostova N. S. Structure and variability of correlations of morphological characteristics of flowering plants: abstract of thesis. diss. … doc. biol. Sciences, 40 (St. Petersburg, 2000).
- Rostova N. S. Correlations: structure and variability, 308(St. Petersburg: St. Petersburg Publishing House. University, 2002).
- Grossheim A. A. Flora of the Caucasus. 2nd ed.. 5: Rosaceae – Leguminosae, 455 (M.L.: Publishing House of the USSR Academy of Sciences, 1952).
- Anatov D. M., Osmanov R. M., Gabibova M. R. Intrapopulation variability of morphological traits of the leaf of Microcerasus incana (Pall.) M. Roem. in Intramountain Dagestan. Izvestiya Gorskogo gosudarstvennogo agrarnogo universiteta, 59(2), 153 (2022).
- Akaev B. A., Ataev Z. V., Gadzhiev B. S., Gadzhieva Z. H., Ganiev M. I. etc. Physical geography of Dagestan 380 (DGPU: Shkola, 1996).
- Mamaev S. A. Forms of intraspecific variability of woody plants, 283 (M.: Nauka, 1972).
- Epskamp S., Cramer A. O., Waldorp L. J., Schmittmann V. D., Borsboom D. qgraph: Network Visualizations of Relationships in Psychometric Data. Journal of Statistical Software, 48(4), 1 (2012).
- Evans J. R., Poorter H. Photosynthetic acclimation of plants to growth irradiance: The relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain. Plant, Cell & Environment, 24, 755 (2001).
- Utkin A. I., Ermolova L. S., Utkina I. A. Surface area of forest plants: essence, parameters, use, 292 (M.: Nauka, 2008).
- Ivanov A. V., Pakomestova V. Y., Kasatkin A. S., Zamolodchikov D. G. Indices of the leaf surface of tree stands in natural and disturbed forests of the Primorsky Territory. Ecology, 4, 243 (2020).
- Niklas K. J. Cobb E. D., Niinemets U., Reich P. B., Sellin A., Shipley B., Wright I. J. “Diminishing returns” in the scaling of functional leaf traits across and within species groups. PNAS, 104, 8891 (2007).
- Filipchenko Y. A. Genetics of soft wheat, 262 (M.-L. OGIZ, 1934).


