
SOME MORPHOMETRIC INDICATORS OF WORKING BEES APIS MELLIFERA LINNAEUS FROM PRIVATE APIERS OF CRIMEA BEEKEEPERS
НЕКОТОРЫЕ МОРФОМЕТРИЧЕСКИЕ ПОКАЗАТЕЛИ РАБОЧИХ ПЧЕЛ APIS MELLIFERA LINNAEUS ИЗ ЧАСТНЫХ ПАСЕК КРЫМСКИХ ПЧЕЛОВОДОВ
JOURNAL: «SCIENTIFIC NOTES OF V.I. VERNADSKY CRIMEAN FEDERAL UNIVERSITY. Biology. Chemistry» Volume 10 (76), №4, 2024
Publication text (PDF):Download
UDK: 57.063
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Bykova T. O., Institute of Biochemical Technologies, Ecology and Pharmacy (structural unit) of the Crimean Federal University named after V. I. Vernadsky, Simferopol, Republic of Crimea, Russia
Lisovaya D. S., Institute of Biochemical Technologies, Ecology and Pharmacy (structural unit) of the Crimean Federal University named after V. I. Vernadsky, Simferopol, Republic of Crimea, Russia
Ivashov A. V., Institute of Biochemical Technologies, Ecology and Pharmacy (structural unit) of the Crimean Federal University named after V. I. Vernadsky, Simferopol, Republic of Crimea, Russia
TYPE:Article
DOI:https://doi.org/10.29039/2413-1725-2024-10-4-26-39
PAGES:from 26 to 39
STATUS:Published
LANGUAGE:Russian
KEYWORDS: honey bee, breed, morphometric characteristics, Crimea.
ABSTRACT (ENGLISH):
Previously, a subspecies of the Crimean bee, described by V. V. Alpatov as A. mellifera taurica (Alpatov, 1948) [4], was widespread in Crimea. However, due to uncontrolled interbreeding and haphazard importation of other races, the subspecies was lost. It is hoped that some families have survived in the foothills or mountain forests of Crimea [5].
In the Crimean peninsula, where for many decades mass imported fetal uteruses of different breeds (subspecies) of European bees, the aboriginal, well adapted to local conditions Taurian breed is now considered lost. A couple of decades ago in natural conditions (mountainous Crimea) [3] bees with a very close phenotype were encountered and there is hope that its descendants can be identified in the process of mass screening of Crimean bee families. In this regard, the main purpose of the study was to compare samples of Crimean bees from four locations in modern times in relation to those bees described as a breed by Prof. V. V. Alpatov in the first half of the last century (1938, 1948). Two apiaries were located in the city of Sevastopol, where reproduction of bee families took place without importation of third-party fertile mothers for more than 20 years; one apiary was located at the edge of the forest on the outskirts of the village of Sokolinoye, where the bee families were reproduced without importation of third-party fertile mothers. One apiary was located at the edge of the forest on the outskirts of Sokolinoye village, where bee generations were reproduced haphazardly by importing fetal mothers from Carpathian, Krainian and other breeds, as well as swarming bees from the adjacent forest.
The last location was a forest glade near Nauchny village, located within a large forest area covering the northern macro-slope of the Crimean Mountains.
We studied 901 specimens of summer collections (June, 2024) of worker honey bees from 25 families from three Crimean apiaries in a forest glade in the mountain-forest zone of Crimea near Nauchny village. In each sample, 20–30 individuals were selected from families and collections in the forest glade and measurements of such most informative morphometric parameters as: proboscis length, cubital index, tarsal index, discoidal displacement were made. The obtained data were processed using Excel 2010 program. The limits of variation and intervals of 95 confidence intervals of arithmetic mean of three quantitative traits and intervals of post-family variation of sample mean proportions of discoidal displacement variants (positive, zero and negative) were determined.
The data of the following authors were used as reference values of the selected indicators: Central Russian (according to V. P. Nikolayenko, 2005) [18], gray mountain Caucasian (according to N. I. Krivtsov, 2009) [19], Carpathian (according to V. A. Gaidar, 2004) [20], Krainian (according to V. V. Alpatov, 1938) [21], Taurian (according to V. V. Alpatov, 1948) [4].
Comparison of these breeds showed that the Taurian bee differs most from the other three southern breeds in cubital index, but at the same time it is noticeably inferior in cubital index value to the Central Russian bee. In discoidal displacement it differs from all breeds by almost complete dominance (94–100 %) of the positive variant.
According to morphometric indices all investigated groups of honey bees from Crimean apiaries do not correspond to any of the most frequently used bee breeds in the territory of the Russian Federation and represent mixed forms, which is caused by non-compliance with breed zoning in Crimea of Carpathian and Ukrainian steppe breeds and the influx of fetal mothers of European subspecies. Worker bees from the families of all four apiaries do not correspond in their morphometric parameters to the reference values of the earlier widely used in Crimea Tauric breed described by Professor V. V. Alpatov of the Moscow State University back in the first half of the last century.
The current Crimean bees studied differ from those in the first half of the last century in the first place by very low values of the cubital index, which is also characteristic of other breeds, as well as by increased frequencies of individuals with negative and neutral discoidal displacement.
In general, modern bees have wider intervals of trait variation, as well as intervals of 95 % confidence intervals of the arithmetic mean, which is caused by the mixing of breeds due to the lack of control over the importation of fetal mothers from European producers.
Thus, this apiary was interesting as a result of the work of an experienced beekeeper for many decades, whose main task was to obtain fertile mothers from the most viable and productive families. So to say mass selection, as it was done by our ancestors for centuries, but with one significant addition: the inclusion in the process of different subspecies originating from European bees as an additional powerful source of replenishment of the gene pool and variability. In fact, even today on the territory of the Crimean peninsula many beekeepers are engaged in this process. The families of this apiary were of interest also for the simple reason that in spring sampling in one family out of 20 analyzed there was one family with identical morphometric parameters of the Tauris breed described long ago. In our summer collection of worker bees from this family there were no more bees with such parameters (polyandry!). Nevertheless, this apiary seemed promising for evaluating the variability of selected traits in the “cauldron” of uncontrolled self-reproduction of bees, in which the gene pool of this apiary was “cooked”, fueled by additives of fetal bee mothers of Carpathian and Krainian breeds popular in the Crimea and, possibly, genes of migratory chicks and swarming families from the adjacent forest (Tauris breed).
ВВЕДЕНИЕ
Медоносная пчела, научное название Apis mellifera Linnaeus, 1758, является частью семейства Apoidea, которое, в свою очередь, объединяет большое количество видов в прогрессирующем отряде перепончатокрылых насекомых Hymenoptera [1].
Пчелы играют важную роль в экосистемах и хозяйственной деятельности человека. Являясь основным опылителем цветковых растений, они выполняют важную функцию в большинстве наземных экосистем. Кроме того, пчелы способствуют плодородию энтомофильных культур. Разводимая человеком медоносная пчела обеспечивает производство продуктов питания и фармацевтических препаратов. Интерес к ним также обусловлен их морфологией и этологией [2].
На территории Крыма еще в советское время были районированы пчелы карпатской (Apis mellifera carpathica Foti) и украинской степной (Apis mellifera sossimai Engel или Apis mellifera acervorum Scor) пород. Сегодня здесь распространены также пчелы кавказской (Apis mellifera caucasica Pollmann), краинской (Apis mellifera carnica Pollmann), итальянской (Apis mellifera ligustica Spinola) пород [3].
Ранее в Крыму был распространён подвид крымской пчелы, описанный В. В. Алпатовым как A. mellifera taurica (Alpatov, 1948) [4]. Однако из-за неконтролируемого межпородного скрещивания и бессистемного ввоза других рас, подвид был утерян. Есть надежда, что некоторые семьи сохранились в предгорьях или горных лесах Крыма [5].
Основным способом классификации признаков пород медоносных пчел считается метод морфометрического анализа. Теоретические основы данного подхода были разработаны А. Г. Кожевниковым (1900, 1906), А. С. Михайловым (1924) и В. В. Алпатовым (1925). Как правило, для характеристики пчел вида A. mellifera применяются такие параметры как: длина хоботка, габариты тела, длина и ширина крыла, кубитальный индекс, дискоидальное смещение, тарзальный индекс и прочие [6].
При морфологических исследованиях пчел наиболее стабильны такие внешние признаки как кубитальный индекс и дискоидальное смещение. Они могут быть применены для определения расовой принадлежности [7, 8]. Проведение таких исследований весьма актуально, так как необходимо получить четкое понимание фенотипического облика крымской популяции A. mellifera, что обеспечит дальнейшую практическую работу по восстановлению ее генофонда и получению высокопроизводительных линий крымской пчелы.
Исторически разные исследования представляют интерес в плане наличия трендов в популяционных дифференциациях. Так Ф. Руттнер более 35 лет назад (1988) идентифицировал 5 популяций Apis mellifera jemenitica из азиатского и африканского регионов (Сомали, Йемен, Судан, Чад, Оман и Саудовская Аравия) и выявил морфометрические различия между популяциями A. m. jemenitica Западной и Восточной Африки. Недавно (2021 г.) в коллективной работе саудовских и египетских исследователей с использованием как морфометрии, так и молекулярно-генетических методов было показано, что популяции медоносных пчел в юго-западном регионе Саудовской Аравии являются уже смешанными. И это явление они увязывают как с завозом линейных пчел A.m. jemenitica, так и с генетической интрогрессией с интродуцированными подвидами медоносных пород [9].
Хорошо известно, что виды могут менять свой географический ареал, внося пластические или адаптивные изменения в ответ на новые условия среды. Фенотипические изменения пчел вида A. mellifera в зависимости от факторов окружающей среды изучались в основном вдоль широтных градиентов, но долгое время оставались относительно мало изученными, несмотря на их важность для экологических взаимодействий [10].
На крымском полуострове, куда на протяжении многих десятков лет массово завозились плодные матки разных пород (подвиды) европейских пчел, аборигенная, хорошо адаптированная к местным условиям окружающей среды таврическая порода, считается ныне утерянной. Еще пару десятилетий в природных условиях [3] встречались пчелы с очень близким фенотипом и есть надежда на то, что ее потомки могут выявиться в процессе массового скрининга семей крымских пчел.
МАТЕРИАЛЫ И МЕТОДЫ
Материалом для исследований послужили особи медоносной пчелы A. mellifera от семей четырех крымских пасек. Первые две группы пчел были собраны на территории двух частных пасек летом в июле 2022 г. в г. Севастополь (Нахимовский район, г. Севастополь, пчеловоды Лисовой С. А. (Рис. 1) и Сагайдачный П. П. (Рис. 2)).

Рис. 1. Ульи с пчелами на первой пасеке в г. Севастополь (фото Лисовой Д. С.)

Рис. 2. Внешний вид второй частной пасеки г. Севастополь (фото Лисовой Д. С.)
Первая пасека действует с 2003 г. на основе плодных маток предположительно карпатской породы, отобранных из второй пасеки, которая существует на протяжении 25 лет, но породность ее пчел до настоящего времени не определялась. Эти пасеки интересны тем, что они располагаются стационарно в городских условиях и воспроизводятся от отводков собственных семей на протяжении более двадцати лет.
Третья группа пчел также неизвестной породности была собрана на цветках нектароносов во время летних сборов в июле 2022 г на лесной поляне, расположенной на юго-восточной окраине поселка Научный (Бахчисарайский район, Республика Крым) (рис. 3).

Рис. 3. Место сбора пчел близ поселка Научный (фото Лисовой Д. С.)
Четвертая группа пчел также неизвестной породности была собрана в июле 2023 г. на территории частной пасеки, расположенной на окраине с. Соколиное (Бахчисарайский р-н, Республика Крым, пчеловод-матковод Спиридонов С. Н.) (рис. 4).

Рис. 4. Общий вид пасеки в с. Соколиное, Бахчисарайский р-н (фото Лисовой Д.С.)
Село расположено в южной части Бахчисарайского района, на окраине леса. Данная пасека существует уже более 15 лет и также не специализировалась на определенной породе. Разведение начиналось с карпатской, краинской и итальянской пород. Самовоспроизводство также происходило за счет собственных отводков; если после зимовки некоторые семьи были слабыми, то в них пчеловод добавлял отводки из других ульев.
Морфометрические показатели остаются по-прежнему востребованными для описания внутривидовой изменчивости. Их комплексы являлся основным критерием классификации, описанные Руттнером (1988) [11]. В настоящем исследовании осуществляли измерения таких показателей как: длина хоботка, кубитальный индекс, тарзальный индекс, дискоидальное смещение. Эти признаки считаются главными, т.к. по ним определяется породная принадлежность пчел на пасеках как в нашей стране, так и за рубежом. Как показал имеющийся опыт многих десятилетий, остальные признаки используются реже, т.к. не дают достаточно высокой разрешающей способности. Например, в недавних исследованиях, проведенных американскими учеными, были продемонстрированы низкие уровни дифференциации между экотипами пчел с использованием традиционной морфометрии их длины [12].
Ниже приводим схему правого переднего крыла медоносной пчелы для более четкого понимания, о каких признаках идет речь (рис. 5). Определение соотношения между длиной межкубитальных жилок “а” и “б” третьей кубитальной ячейки переднего крыла, выраженное в процентном отношении является кубитальным индексом.
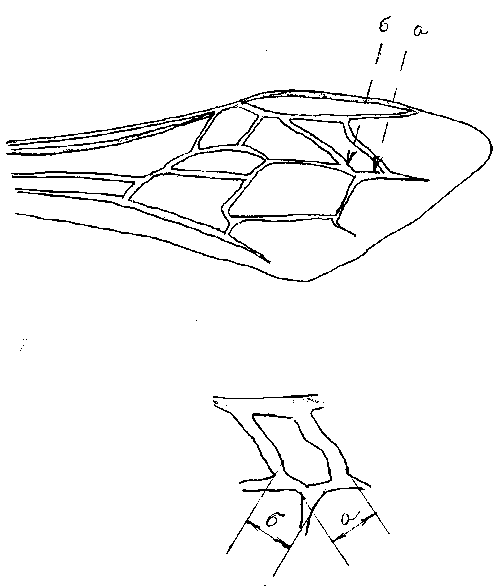
Рис. 5. Правое переднее крыло пчелы медоносной [4, 7]
а – длина межкубитальной жилки третьей кубитальной ячейки,
б – длина межкубитальной жилки.
Следует отметить, что длину хоботка как функционально более важную характеристику приспособленности к сбору нектара у нектароносов с различной глубиной залегания нектара можно применять для дифференциации пасек [13].
Вместе с измерениями длины хоботка по-прежнему в качестве наиболее объективных признаков используют кубитальный и тарзальный индексы [14]. Привлечённые нами показатели входят в пул более широкого перечня, но
по-прежнему являются одними из наиболее информативных, что, например, недавно было показано корейскими учеными при определении комплекса морфометрических признаков у местных адаптированных линий Apis cerana koreana, A. melifera ligustica, A. mellifera carpathica) [15].
После сопоставления границ вариации референсных значений морфометрических показателей у пчел различных пород, распространенных на территории Российской Федерации, с полученными в прошлом веке по таврической пчеле A. mellifera taurica (Alpatov, 1948) [4], проводили сопоставления наших результатов с данными по крымским пчелам, исследованными в нынешнее время [16, 17].
Результаты измерений осуществляли с использованием окуляр-микрометра и бинокуляра МБС-9. Данные обрабатывались с привлечением статистических возможностей программы Excel. При оценке достоверности различий конечные показатели подавались в виде интервалов доверительной вероятности, вычисленных на 5 %-ном уровне значимости. Для этого стандартные ошибки средней арифметической умножали на табличный t0.05— критерий Стьюдента, найденный для числа степеней свободы f = n-1 и уровня доверительной вероятности 95 %.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Для сравнения результатов изучения крымских пчел с опубликованными данными по другим породам, в том числе полученными в тридцатые–семидесятые годы прошлого столетия, когда статистическая обработка материалов проводилась далеко не всегда, применяли оценку показателей в виде границ вариации. Таким образом, получили возможность использовать референсные данные, опубликованные в известных классических исследованиях, которые были выполнены в прошлом веке, и сопоставлять их с современными.
По данным таблицы 1, можно увидеть достаточно четкие различия границ вариации референсных значений морфометрических показателей карпатской, среднерусской, серой горной кавказской, краинской и таврической пород. К сожалению, данных по границам вариации кубитального и тарзального индексов у таврической пчелы в публикациях В. В. Алпатова мы не нашли и поэтому вставили интервал доверительной вероятности.
По длине хоботка таврическая пчела не выделяется среди других пород. Породный стандарт только серой горной кавказской породы медоносной пчелы отличается от других самым широким и большим интервалом варьирования длины этого показателя. Обращает на себя внимание наименьшее значение длины хоботка у среднерусской породы, что не противоречит правилу Аллена [11], согласно которому в более прохладном климате длина хоботка уменьшается. Скорее всего, это является результатом естественного отбора, обусловленного глубиной залегания нектара в цветках липы сердцевидной, как одного из основных кормовых растений медоносной пчелы на обширных территориях Евразии [22–25].
Таблица 1
Границы вариации референсных значений морфометрических показателей у различных пород медоносных пчёл, наиболее часто используемые на территории Российской Федерации
|
Границы вариации морфометрических показателей |
||||||
|
Длина хоботка, мм |
Кубиталь-ный индекс, % |
Тарзальный индекс, % |
Дискоидальное смещение, % |
|||
|
«+» |
«0» |
«–» |
||||
|
Среднерусская (по Николаенко В. П., 2005) [18] |
6,0–6,4 |
60–65 |
55–60 |
0 |
5–10 |
90–95 |
|
Серая горная кавказская (по Н. И. Кривцову, 2009)[19] |
6,7–7,2 |
50–55 |
50–60 |
0 |
0–29 |
71–96 |
|
Карпатская (по В. А. Гайдару, 2004) [20] |
6,6–6,7 |
33–43 |
57–60 |
85–100 |
0–15 |
0–5 |
|
Краинская (по В. В. Алпатову, 1938) [21] |
6,4–6,8 |
46–47 |
58–61 |
80–100 |
0–20 |
0–5 |
|
Таврическая (по |
6,4–7,2 |
57,98±2,24 |
57,73±0,38 |
94–100 |
0–6 |
0 |
По кубитальному индексу таврическая пчела отличается значительно большими значениями минимальных и максимальных величин среди четырех южных пород, но, в то же время, заметно уступает по величине кубитального индекса среднерусской.
Интервалы значений тарзального индекса очень близки к референсным значениям пчел всех пяти пород. Характерно, что максимальные величины (верхние границы) у всех пород практически совпадают, но нижние границы – нет. В этом случае можно отметить наименьшее значение у серой горной кавказской породы. Таким образом, тарзальный индекс менее информативен при различении подвидов, чем кубитальный. И это важно учитывать при поиске и восстановлении таврической пчелы.
По доминированию отрицательного дискоидального смещения среднерусская и серая горная кавказская породы пчел кардинально отличаются от остальных пород. Однако, для всех породных групп свойственно наличие небольшой доли пчел с нейтральным дискоидальным смещением. При этом таврическая пчела в прошлом веке отличалась от всех остальных наибольшей «чистотой» по этому признаку, своим почти 100 %-ным доминированием положительного дискоидального смещения. Это также важно учитывать при поиске семей этой пчелы с такими характеристиками.
В таблице 2 представлены современные границы вариации морфометрических показателей у пчел из четырех крымских пасек в сопоставлении с ранее распространенной здесь таврической пчелой.
Таблица 2
Средние значения, интервалы их доверительной вероятности и границы вариабельности морфометрических показателей у современных крымских пчел и пчел таврической породы A. mellifera taurica (Alpatov, 1948) [4]
|
Группа пчел |
Интервалы доверительной вероятности 95 % и границы вариации морфометрических показателей |
|||||
|
Длина хоботка, мм x̅±∆ Lim |
Куб. индекс, % x̅±∆ Lim |
Тарзальный индекс, % x̅±∆ Lim |
Дискоидальное смещение, Lim % |
|||
|
«+» |
«0» |
«-» |
||||
|
Пчелы первой частной пасеки г. Севастополя от 3 семей (n=113) |
6,23±0,18 5,4–7,2 |
41,43±2,24 26,8–58,3 |
58,11±0,41 47,6–64,1 |
32–100 |
0–44 |
0–42 |
|
Пчелы второй частной пасеки из г. Севастополя от 5 семей (n=142) |
6,4±0,15 5,3–7,7 |
40,87±2,13 27,3–58,8 |
58,97±0,65 53,5–62,8 |
24–77 |
13–48 |
4–28 |
|
Пчелы частной пасеки из п. Научный собраны на цветках (n=25) |
6,00±0,23 5,2–6,9 |
41,45±2,22 32,6–56,4 |
56,66±0,97 52,4–61,0 |
56* |
36* |
8* |
|
Пчелы частной пасеки из с. Соколиное от 20 семей (n=621) |
6,56±0,08 4,7–7,0 |
41,57±1,94 24,0–64,0 |
57,82±0,65 52,3–65,9 |
0–100 |
0–52 |
0–42 |
|
Таврическая пчела от 3 семей (n=621) (по В. В. Алпатову, 1948) [4] |
6,44±0,06 |
57,98±2,24 |
57,73±0,38 |
94–100 |
0–6 |
0 |
Примечание: *В пос. Научный исследовалась выборка из пчел, кормившихся на лесной поляне, поэтому интервалы варьирования значений дискоидального смещения не представлены.
Как видно из представленных данных, интервалы доверительной вероятности и границы интервалов варьирования у всех исследуемых пчел из крымских пасек по всем проанализированным показателям, за исключением кубитального индекса, шире, чем у таврической породы.
Таким образом, на сегодняшний день у 75 % исследованных крымских пчел интервалы варьирования всех количественных признаков, также, как и у качественного признака (дискоидальное смещение), гораздо шире. При этом, как видно из табличных данных, появилась большая доля особей во всех четырех изученных группах пчел с нейтральным вариантом дискоидального смещения, чего не наблюдалось в Крыму в прошлом столетии. Конечно, в условиях несоблюдения поддержания чистоты пород, районированных на полуострове еще в советское время, отсутствие современной стратегии развития пчеловодства на данной территории, массовая интродукция других пород из европейских стран, этого и следовало ожидать. Как известно, аналогичные проблемы существуют с поддержанием чистоты коренных популяций среднерусской пчелы, о чем имеется достаточно актуальных свидетельств [26–28].
Попарное сравнение интервалов доверительной вероятности количественных данных, полученных для крымских пчел из 4-х пасек с ранее существовавшей здесь таврической породы, показало, что все эти пчелы по тарзальному индексу близки и даже с привлечением статистической обработки, не имеют достоверных различий с таврической пчелой. Все нынешние крымские пчелы имеют значения кубитального индекса ниже, чем у таврической.
По длине хоботка все анализируемые группы пчёл отличаются от таковых у A. mellifera taurica более низкими значениями данного параметра. Этот показатель достаточно часто используется для дифференциации пчелиных семей на пасеках. Так, например, недавно (август, 2024г.) морфометрические характеристики A. mellifera в Суданской и Судано-Сахельской климатических зонах Африки (800 рабочих пчел из 80 семей 16 пасек Суданской и Судано-Сахельской климатических зон Буркина Фасо) показали также достаточно широкий интервал доверительной вероятности длины хоботка (4,31±0,57 мм). Это доказывает, что интродуцированная западная медоносная пчела на другом континенте стала иметь значительное меньшее значение длины хоботка и большую вариабельность этого показателя [29].
Как свидетельствуют данные таблицы 1, в прошлом у таврической породы, как и у карпатской и краинской, преобладало положительное дискоидальное смещение, но если у таврической не встречались особи с отрицательным, и очень редко с нейтральным смещением, то у двух других – эти качественные признаки достигали заметных значений. На сегодня у крымских пчел выявлены особи с нейтральным и отрицательным дискоидальным смещением в значительном количестве, что не было отмечено в прошлом.
ЗАКЛЮЧЕНИЕ
- По морфометрическим показателям все исследуемые группы медоносных пчел из крымских пасек не соответствует ни одной, из наиболее часто используемых на территории Российской Федерации породе пчел, и представляют собой помесные формы, что обусловлено несоблюдением породного районирования в Крыму карпатской и украинской степной пород.
- Рабочие пчелы от семей из всех четырех пасек по своим морфометрическим показателям не соответствуют референсным значениям ранее широко разводимой в Крыму таврической породе, описанной профессором МГУ Алпатовым В. В. еще в первой половине прошлого века.
- Исследованные крымские пчелы отличаются в первую очередь очень низкими значениями кубитального индекса, что свойственно для других пород, а также возросшими значениями частот встречаемости особей с отрицательным и нейтральным дискоидальным смещением.
- Современные пчелы имеют более широкие интервалы варьирования признаков, также как и интервалы 95 %-ной доверительной вероятности средней арифметической, что вызвано смешением пород из-за отсутствия контроля ввоза плодных маток от европейских производителей.
- L. Ya. Moreva Ecological features of the honey bee (Apis mellifera L.) in the south of Russia, Abstract of a dissertation for the degree of Doctor of Biological Sciences, 48 p. (Moscow, 2007).
- Cherevko Yu. A., Cherevko L. D., Boytsenyuk L. I., Kochetov A. S. A. Beekeeping: a textbook for universities on agronomic specialties , 296 p. (Moscow, Kolos, 2008).
- Ivashov A. V., Bykova T. O., Sattarov V. N., Mannapov A. G. Condition and preservation of Apis mellifera taurica in the Crimea, Journal “Beekeeping”, 9, 20 (2016).
- Alpatov V. V. Honey bee breeds, 183 p. (Publishing House of the Moscow Society of Nature Testers, (1948).
- Ruttner F. P. Breeds of bees in Africa, XXV International Congress on Beekeeping, 330 (1976).
- Berezin A. S. Methods of morphometry in determining the breed of honey bees, Biomics, 11(2), 167 (2019).
- Lavrekhin F. A., Pankova S. V. Biology of the honey bee, 303 p. (Kolos, Moscow, 1983).
- Mărghitaş A. L., Paniti-Teleky O., Dezmirean D. S., Mărgăoan R., Bojan C., Coroian C. L., Moise L. A., Morphometric differences between honey bees (Apis mellifera carpatica) populations from Transylvanian area, Journal Scientific Papers Animal Science and Biotechnologies, 41, 300 (2008).
- Enas A. A., Alabdali H., Ghramh, A., Essam H., Ibrahim B., Zubair A., Asma N. Asiri of the native honey bee (Apis mellifera jemenitica) in the south western region of Saudi Arabia using morphometric and genetic (mtDNA COI) characteristics . Saudi J Biol Sci., 28(4), 2278 (2021).
- Gérard M., Martinet B., Maebe K., Smagghe G., Vereecken N. J., Rasmont P. Shift in size of bumblebee queens over the last century. Glob. Change Biol., 26, 1185 (2020).
- Ruttner F. Biogeography and taxonomy of honey bees, 284 p. (Berlin, Germany: Springer-Verlag, 1988).
- Calfee E., Agra M. N., Palacio M. A., Ramírez S. R., Coop G. Selection and hybridization shaped the rapid spread of African honey bee ancestry in the Americas, e-print arXiv: e1009038 (2020).
- Bianchi E., Agra M. N., García C., Gennari G., Maldonado L., Rodríguez G. A., Palacio M. A., Scannapieco A. C. and Lanzavecchia S.B. Defensive Behavior and Morphometric Variation in Apis mellifera Colonies From Two Different Agro-Ecological Zones of North-Western Argentina, e-print arXiv: 590225 (2021).
- Kakhramanov B., Isamukhamedov S., Safarova F., Donaev K., Mamadov F. Morphological features of the Carniolan (Apis mellifera carnica Pollm) and Carpathian (Apis mellifera carpatica) breeds of honey bees, e-print arXiv: 03018E3 (2021).
- Frunze O., Сhoi Y. S., Kim D. W., Park B. S., Park H. G., Kang E. J. Morphometric Characterization of Honey Bee, Apis mellifera Linnaeus, Inbred Lines in Korea. Korean Journal of Applied Entomology, 59 (4), 371( 2020).
- Bykova T. O., Krivozubov A. S., Ivashov A. V., Sattarov V. N., Safonkin A. F., Triseleva T. A., Soloviev A. M., Emirsinov I. S. Morphometric variability of wild honey bees of the mountain forest zone of Crimea as a material for breeding, e-print arXiv: 04014 (2020).
- Sattarov V. N., Skvortsov A. I., Ivashov A. V., Bykova T. O., Ivanov S. P., Emirsinov I. S., Mishchuk R. M., Sabirjonova M. R., Kistanova R. R., Sharafutdinov D. Z. Epidological collections, Magazine “Beekeeping”, 7, 10, (2020).
- Nikolaenko V. P. Breeding work with bees, 144 p. (BARO-PRESS, Rostov, 2005).
- Krivtsov N. I., Sokolsky S. S., Lyubimov E. M. Gray mountain Caucasian bees, 192p. (Scientific publication, Sochi, 2009).
- Gaidar V. A. Comparative assessment of Carpathian and Krajin bees, Beekeeping, 8, 18 (2004).
- Alpatov V. V. Bees of the Crimea and the Crimea and their place among other forms, Zool. journal., 3, 472 (1938).
- Bykova T. O., Ivashov A. V., Sattarov V. N., Amelina V. O., Sattarova A. V. Assessment of bee colonies in the steppe zone of Crimea, “Beekeeping”, 10, 23 (2021).
- Ruttner F. Biometrical-statistical analysis of the geographic variability of Apis mellifera L, Apidologie, 9 (4), 363 (1978).
- Ruttner F. Naturgeschichte der Honigbienen, 455 p. (Munich: Ehrenwirth, 1992).
- Krivtsov N. I. Identification of intraspecific taxa of honey bee, Beekeeping, 8, 23 (2003).
- Sagitov S. T., Sattarov V. N., Abdrakhimova Y. R., Zainullina G. R., Sultanova R. R., Khannanova L. F. Study of the Apis mellifera population in the territory of the Toratau Geopark as a search for an additional reserve of purebred bees, Bulletin of M. Akmulla Bashkir State Pedagogical University, №S1 (66) (2023).
- Sagitov S., Ilyasov R., Sattarov V., Abdrakhimova Y., Danilenko V., Gazizova N., Sattarova A., Boguslavsky D. Phenetic diversity of the honey bee population (Apis mellifera) in the Toratau geopark of the Republic of Bashkortostan, U. Bee J., 23(1), 115 (2023).
- Sabirdjhonova M. R., Sattarov V. The phenotypic variability of Apis mellifera drones in apiaries of the northern part of Bashkortostan, Izvestiya vysshikh uchebnykh zavedeniy. Povolzhskiy region. Estestvennye nauki = University proceedings. Volga region. Natural sciences, 2, 74 (2021).
- Souhaïbou S., Hugues R. B., Oswald G. D. Morphometric characteristics of the honeybees (Apis mellifera Linnaeus, 1758) and their variability in the Sudano-Sahelian and Sudanian climatic zones of Burkina Faso, Authorea (2024).