
ONTOMORPHOGENESIS AND CОENOPOPULATION STRUCTURE OF CREPIS PURPUREA (WILLD.) M. BIEB. IN CRIMEA
JOURNAL: «SCIENTIFIC NOTES OF V.I. VERNADSKY CRIMEAN FEDERAL UNIVERSITY. Biology. Chemistry» Volume 6 (72), №3, 2020
Publication text (PDF):Download
UDK: 574.34(58.009)
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Abdulganieva E. F., V. I. Vernadsky Crimean Federal University, Simferopol, Russia
TYPE:Article
DOI:https://doi.org/10.37279/2413-1725-2020-6-3-3-17
PAGES:from 3 to 17
STATUS:Published
LANGUAGE:Russian
KEYWORDS: Crepis purpurea (Willd.) M. Bieb., ontomorphogenesis, ontogenetic structure, vitality, spatial structure.
ABSTRACT (ENGLISH):
The study is devoted to ontomorphogenesis and the structure of cоenopopulations of the rare species Crepis purpurea (Willd.) M. Bieb., protected by the Red Data Book of Sevastopol, the Republic of Crimea, Turkey and I Appendix of the Berne Convention.
The duration, morphological and anatomical features of the age states in the pregenerative, generative and senile periods of C. purpurea ontogenesis are established. The total duration of ontogenesis is 5–6 years, which includes 4 ontogenetic periods and 9 age states. C. purpurea is characterized by the structural type of polyvariance with the formation of 3 ontobiomorphic variants: individuals with one-, two-, or many-headed caudex. The features and the period of the onset of anatomical partition in the shoot part caudex were revealed. Germination of C. purpurea seeds was determined.
The ontogenetic, vitality and spatial structure of 11 Crimean cоenopopulations of C. purpurea were carried out. The lack of the pregenerative and senile individuals fractions is observed in coenopopulations. The basic ontogenetic spectrum of the studied populations is bimodal with a peak on virgin and mature generative individuals. The indices of renewability, generativeness, aging, the general age of the population, as well as the age indices of A. A. Uranov and the efficiency of L. A. Zhivotovsky were calculated for each population. In the vital structures there is a dynamic on the increase in the values of the index of vital quality of populations from the eastern foothills to the western foothills and further – to the mountain plateaus variants of populations. It was established that individuals of C. purpurea have a tendency to tend to a group type of distribution in the population. The data on areas of coenopopulations, the number and density of individuals of C. purpurea in the studied communities are presented.
ВВЕДЕНИЕ
Эффективное сохранение редких и исчезающих видов, в первую очередь, зависит от научно обоснованных охранных мер, направленных на их защиту на популяционном уровне. В связи с этим, особенно важным является знание структуры популяций и особенностей жизненного цикла, без понимания которых невозможна объективная интерпретация результатов оценки современного состояния и прогнозирование популяционной динамики видов [1]. Наиболее информативным в этом смысле для растительных видов являются анализ онтогенетической, виталитетной и пространственной структур популяций.
Одним из компонентов сообществ, произрастающих на обнажениях известняковых и мергелистых пород Горного Крыма, является редкий Crepis purpurea (Willd.) M. Bieb. Скерда пурпуровая имеет ограниченный в мире ареал (Крым, Северная Турция), большая часть популяций которого сохранилась на территории Крымского полуострова [2, 3], что послужило обоснованием необходимости включения вида в региональные Красные книги города Севастополя [4], Крыма [5] и Турции [6], а также в международный Красный список растений, охраняемых Бернской конвенцией [7]. Исследований, охватывающих комплексное изучение характеристик предгорных и яйлинских вариантов популяций, ранее не проводилось.
Целью данной работы является изучение особенностей онтоморфогенеза и структуры ценопопуляций (ЦП) C. purpurea в Крыму.
МАТЕРИАЛЫ И МЕТОДЫ
Исследования проводились в течение вегетационных сезонов 2018–2019 годов в 11 фитоценозах Главной и Внутренней гряд Крымских гор, в составе которых произрастает C. purpurea (рис. 1).
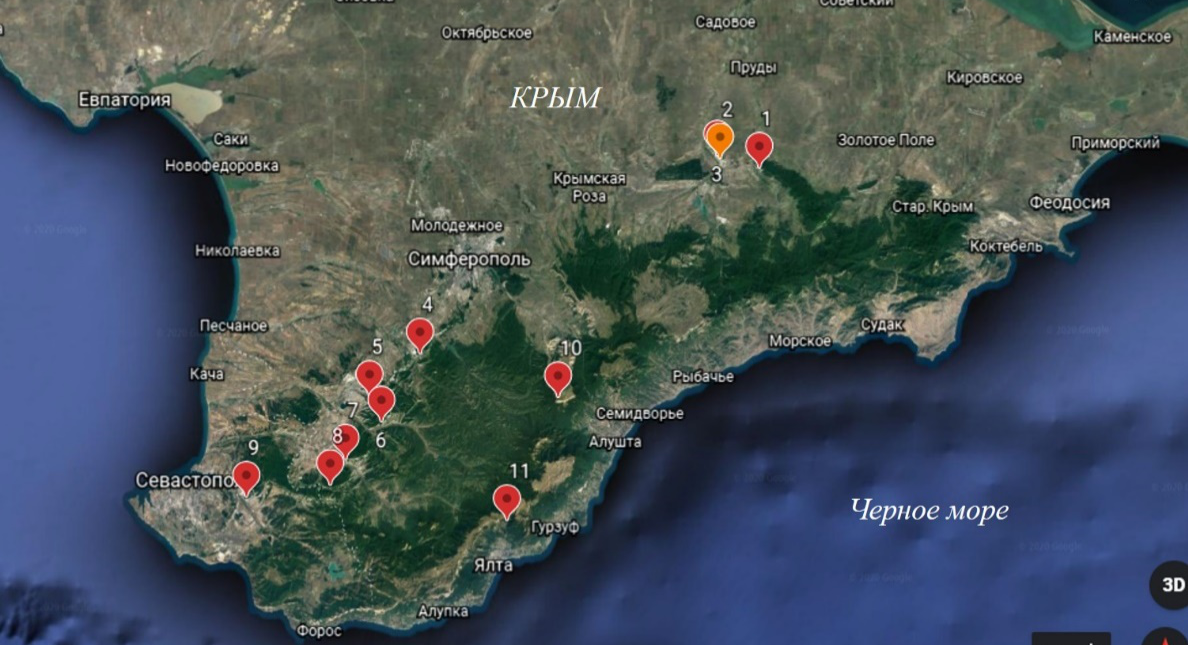
Рис. 1. Карта расположения исследуемых ценопопуляций C. purpurea.
Дифференциация и характеристика возрастных состояний C. purpurea базировались на концепции дискретного описания онтогенеза, предложенной Т. А. Работновым [8] и детально разработанной его последователями А. А. Урановым [9], О. В. Смирновой [10] и Ю. А. Злобиным [11]. Определение типа онтогенеза и вариантов поливариантного развития проводилось по классификациям Л. А. Жуковой [12–15]. Онтогенетическая структура ценопопуляций анализировалась с привлечением онтогенетических индексов И. Н. Коваленко [16]: возобновляемости (Iв), генеративности (Iг), старения (Ic), общей возрастности популяции (I возраст), а также индексов возрастности (Δ) А. А. Уранова [9] и эффективности (ω) Л. А. Животовского [17]. Виталитетная структура ЦП устанавливалась по состоянию 30 средневозрастных генеративных особей согласно методике Ю. А. Злобина [1, 18] в программе «VITAL». Для определения характера пространственного распределения особей использовался индекс Одума (IO), [19], в качестве размера счетной единицы площади был принят 1 м2.
Названия видов даны по [20].
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Crepis purpurea (Willd.) M. Bieb. [Lagoseris purpurea (Willd.) Boiss.; L. callicephala Juz.] – многолетний травянистый стержнекорневой поликарпик семейства Asteraceae [4, 21], (рис. 2).

Рис. 2. а – Генеративная особь Crepis purpurea; b – Участок ценопопуляции № 7 (Бельбекская долина, подножье скалы Топчи-Кая).
Существование морфологически близких видов L. purpurea, L. callicephala и L. robusta Czer., основанием для выделения которых, главным образом, являются различия в характере опушения листочков обертки корзинки и цветоносов, до сих пор остается спорным и не имеет единого решения [22, 23]. Так, в современной литературе онтогенезу и возрастной структуре 2-х популяций L. callicephala посвящена работа А. Р. Никифорова [24]. Автором также отмечаются фенологические различия в развитии побеговых систем L. callicephala и L. purpurea [25]. Однако в последней флористической сводке высших растений Крыма [20] указывается, что вышеуказанные таксоны не имеют статуса вида и отнесены в разряд синонимов C. purpurea.
В виду вышесказанного, а также учитывая, что в составе исследованных сообществ присутствовали особи с разным типом опушения корзинок и цветоносов, изученные ценопопуляции рассматривались в объеме одного вида – C. purpurea.
В результате морфолого-анатомического анализа в жизненном цикле C. purpurea были выделены 4 онтогенетических периода (латентный, прегенеративный, генеративный, постгенеративный) и 9 возрастных состояний.
Латентный период представлен семенами (se). Плод – семянка, вытянута в длину до 3–6 мм, от черного до темно-коричного цвета, со слабозаметными ребрами. Распространению семян способствует хорошо развитый паппус грязно-белого цвета, почти одинаковой длины с семенем – 2–5 мм длины.
Прегенеративный период. Проростки (p) Наблюдение за проростками частично было проведено в лабораторных условиях. Прорастание семян происходит на 14 сутки при температуре 20 ○С. Всхожесть семян низкая, так как составляет всего 11 %. Прорастание семян надземное. Проростки с 2 простыми семядольными линейными листьями: длиной 6–13 мм, шириной 1–2 мм. Листовые пластинки следующей пары листьев p остаются цельными, но форма становится лопатчатой. Гипокотиль различной длины, и зависит от особенностей микрорельефа местообитания. Появление проростков в природе приурочено к середине весны – началу лета. Переход в следующее возрастное состояние происходит через 1–1,5 месяца.
Особенности развития ювенильных растений (j) определяют дальнейший ход онтоморфогенеза. В типичном случае растение в этом возрастном состоянии представляет собой первичный побег 2–2,5 см высотой, имеющий моноподиальное нарастание и 3–4 листа обратно-узкояйцевидной формы с 1–2 парами зубчиков, расположенными ближе к верхушке листовой пластинки (13–18 мм длины и 3–5 мм ширины). Главный корень удлиняется до 5–8 см и развивает нитеобразные боковые корни. Переход в следующее возрастное состояние происходит через 1 месяц. В случае гибели верхушечной почки (заморозки, засыпание почвой, что случается часто на этих субстратах) происходит развитие пазушных почек со сменой моноподиального нарастания на симподиальное. В зависимости от развития числа побегов замещения, в дальнейшем будет формироваться симподиально нарастающая стержнекорневая биоморфа с одно-, дву- или многоглавым каудексом. Поливариантность развития ювенильных растений демонстрирует пластичность онтогенеза и обеспечивает более высокие шансы выживания наиболее уязвимой прегенеративной группы популяции. Похожие формы полиморфизма ювенильных и имматурных особей были также выявлены и у других каудексовых растений Scorzonera glabra Rupr. [26], Кnautia arvensis (L.) Coult., Helichrysum arenarium (L). Moench. [27] и Jurinea cyanoides [28].
Переход в имматурное (im) возрастное состояние происходит к концу-середине лета. Имматурные особи формируют розеточный побег с диаметром розетки 5–6 см (по длине самого длинного листа 2,5–3 см). Листорасположение листьев из супротивного переходит в очередное. В розетке развивается 4–6 полувзрослых листьев, имеющих перисто-раздельную листовую пластинку, у которой крупные доли листа чередуются с короткими зубцами. Основание главного корня утолщается (до 2 мм), одновременно продолжается нарастание главного и боковых корней. В этом состоянии растение находится 1–1,5 месяца.
Виргинильные особи (v) – стержнекорневые, однопобеговые, высотой 4–5 см. Количество листьев в розетке 4–6. Форма и размеры листьев достигают размеров листьев взрослой особи (длина 17–90 мм; ширина 4–60 мм). Листья взрослого типа дважды перисто-раздельные, с мелкими зубцами между крупными долями листа. В результате утолщения гипокотиля и основания главного корня формируется каудекс (3–4 мм диаметром), от его корневой части отходят боковые корни по толщине равные главному корню, который к этому времени уже начинает замедляться в росте. Каудекс, достигающий 1 см в длину, втягивается в почву. В этом возрастном состоянии растение уходит на первую перезимовку. По нашему мнению, вид проявляет экологическую пластичность и в разных климатических условиях ведет себя как летне-зимнезеленое – в Предгорье, и как летнезеленое растение – в популяциях, произрастающих на яйлах Крымских гор.
Генеративный период. К цветению способны растения 2-го года жизни. У молодых генеративных (g1) растений розеточный побег образует генеративные побеги максимум с 1–3 корзинками. Первым зацветает терминальный генеративный побег, затем к цветению приступают пазушные побеги. В типичном случае каудекс одноглавый. При развитии дву- (или многоглавого) каудекса каждый из них проходит индивидуальный путь развития. Всего развивается 1–3 флоральных побега, которые к концу вегетационного сезона отмирают, еще год сохраняясь на материнской особи. Количество листьев в розетке от 5–6 (7). Высота растения по генеративному побегу 5–12 см. Прирост длины каудекса за один год составляет около 1 см, его диаметр в этом возрастном состоянии 3–5 мм.
Средневозрастные генеративные растения (g2) достигают максимального развития. Высота растения 15–35 см. На каудексе развиваются боковые розеточные побеги, функционирующие 1–2 года. После цветения базальные части розеточных побегов входят в состав многолетней структуры каудекса, в результате чего каудекс становится многоосевым, в связи с чем провести четкие морфологические границы между разными вариантами онтобиоморф затруднено. В анатомическом строении побеговой части каудекса (корневища) проявляются признаки партикуляции, распространяющейся в центробежном направлении, что приводит к образованию центральной полости (рис. 3).
Длина каудекса 3–4 см, диаметр 5–7 мм. При благоприятных условиях боковые розеточные побеги способны развивать генеративные побеги уже в первый год своего развития. Количество генеративных побегов, образующихся за один вегетационный сезон, достигает 4–7. В случае многоглавого каудекса количество цветущих побегов значительно увеличивается (наблюдаемый максимум – 16 шт.). Число корзинок на одном генеративном побеге 3–7. При горизонтальном расположении каудекса в субгоризонтальном пространстве способны развиваться придаточные корни. Длительность пребывания в средневозрастном генеративном состоянии 1–2 (3) года.
Для старых генеративных растений (g3) характерно снижение интенсивности цветения. Число генеративных побегов уменьшается, они становятся схожими по структуре и высоте с молодыми генеративными особями. Высота растения колеблется от 10 до 15 см. Количество цветоносных побегов на одном каудексе 1–3. Каудекс покрывается мощным слоем перидермы и отмершими основаниями листьев, достигая толщины 8–15 мм. Структура каудекса становится очень рыхлой, он легко распадается при попытке выкапывания, верхние ответвления каудекса (каудикулы в понимании Федорова и др., 1962) отмирают. Процессы партикуляции усиливаются, что особенно заметно у особей с многоглавыми каудексами, у которых распад тканей захватывает как побеговую, так и корневую части каудекса. Новых боковых розеточных побегов закладывается очень мало (1–2). В этом состоянии растение находится 1 год. Общая длительность генеративного периода 3–4 года.
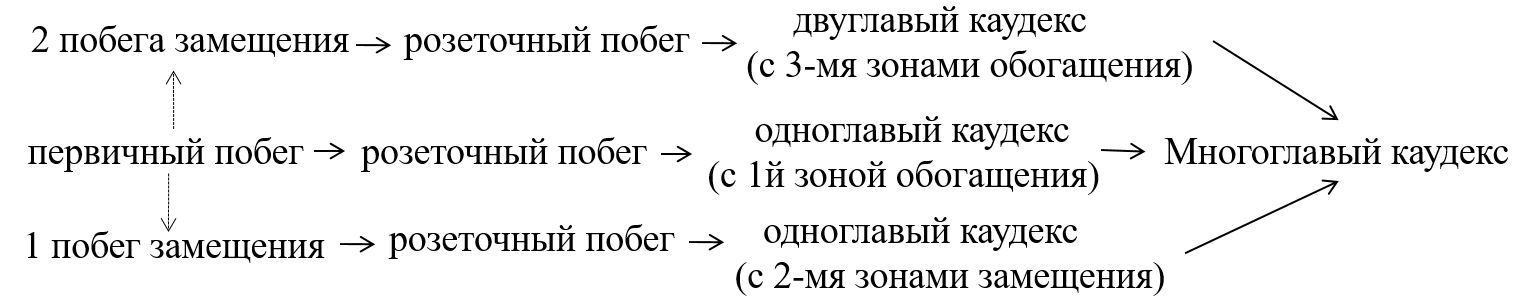
Рис. 3. Партикуляция в побеговой части каудекса (корневища) Crepis purpurea.
Постгенеративный период. До субсенильного возрастного состояния (ss) растения доживают редко и, не проходя заметного сенильного периода (s), как правило, выпадают из популяции в конце позднего генеративного возраста. Сенильные особи представлены остатками распадающегося каудекса, с несколькими малоразвитыми розеточными побегами и «полувзрослыми» (имматурными) листьями. В некоторых случаях при распаде каудекса розеточные побеги, имеющие придаточные корни, способны к самостоятельному существованию еще 1–2 года. Продолжительность постгенеративного периода 1 год.
Для особей C. purpurea характерен структурный тип поливариантности, при котором формируется 3 варианта онтобиоморфы – особи с одно-, дву- или многоглавым каудексом, которые к g3 возрасту становятся трудно различимы без детального морфологического изучения, проходя следующие стадии развития (рис. 4).
Рис. 4. Стадии развития Crepis purpurea.
Общая продолжительность жизненного цикла C. purpurea в среднем составляет 5–6 лет. В классификации основных типов онтогенезов растений онтогенез C. purpurea относится к А–типу и подтипу А 2, при котором весь жизненный цикл проходит одна вегетативно неподвижная особь в течение 3–30 (и более, если благоприятные условия) лет с осуществлением и постгенеративного периода. Таким образом, C. purpurea по типу биоморфы является слабо партикулирующим стержнекорневым поликарпиком (рис. 5).

Рис. 5. Онтоморфогенез особей Crepis purpurea.
Онтоморфогенез особей C. purpurea. 1 – типичный вариант развития j особей; 2, 3 – поливариантные типы развития j особей; а – имматурный лист;
b – виргинильный лист.
На основании знаний о ходе онтоморфогенеза C. purpurea была изучена онтогенетическая структура исследуемых популяций. Учет возрастных состояний проводился на пробной площади 100 м2 в связи с неравностью площадей природных популяций. Общее число особей в популяциях колеблалось от 9 до более 1000 особей. Основная роль в сложении популяций C. purpurea принадлежит прегенеративной (виргинильные особи) и генеративной фракциям – (средневозрастные генеративные особи). Из 11 исследуемых популяций только 3 являются полночленными (ЦП 2, 3, 4). Другие 8 популяций – неполночленные с выпадением прегенеративных (ЦП 10) и сенильных особей (ЦП 1, 5, 7) или обеих групп (ЦП 6, 8, 9, 11). Отсутствие в популяции растений прегенеративной фракции свидетельствует о нарушении процессов возобновления ЦП в определенных сообществах, что, в свою очередь, зависит как от качества и количества семенных зачатков в популяции, так и условий среды обитания.
Из исследованных 200 семян доля нормально развитых семян достаточно высока – 75 %. Однако исследование всхожести семян показало очень низкие значения (11 %) этого показателя для C. purpurea. Важную роль также играют особенности микрорельефа местности, в которой произрастают популяции. Так, например, с повышением угла крутизны склона существенно снижается как общее число особей в популяции, так и участие прегенеративной группы. Кроме того, аэрохорный тип распространения семян может играть негативную роль, в результате которого семена уносятся за пределы популяции и попадают в сообщества, где возможность их развития лимитирована. Таким образом, минимальная площадь популяций, соответствующая понятию «элементарная демографическая единица» [29], в которых обнаруживаются все характерные для C. purpurea онтогенетические группы, находятся в пределах 143–470 м2.
Базовый онтогенетический спектр изученных популяций C. purpurea бимодальный с пиком на виргинильных (v) и среднегенеративных особях (g3), что указывает на то, что, несмотря на сложности появления и неравномерность выживания проростков в разные годы, в популяциях приходит достаточное накопление прегенеративной фракции (рис. 6).
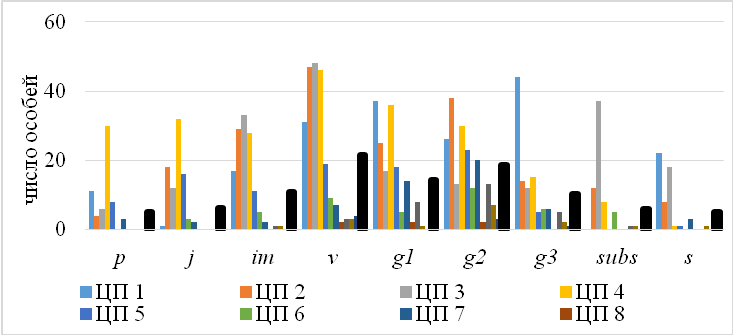
Рис. 6. Онтогенетические спектры популяций Crepis purpurea.
На основе соотношения онтогенетических состояний для популяций были рассчитаны онтогенетические индексы: индекс восстановления (Iв), индекс старости (Iс), индекс генеративности (Iг), индекс возрастности (Iвозраст.), коэффициент возрастности (Δ) и индекс эффективности (ω).
Согласно И. Н. Коваленко, при значениях индекса возрастности (Iвозраст.) меньше 1 – в популяциях преобладают процессы деградации, больше 1 – преобладают инвазионные процессы. Таким образом, к первой группе ЦП C. purpurea относятся ЦП 1, 9 и 10 (Iвозраст. – 1,10; 1,00 и 1,50 соответственно), ко второй – принадлежит большинство ЦП (ЦП 2–8, 11), у которых значения Iвозраст колеблется в пределах 0,00–0,68. В первую очередь низкие значения Iвозраст указывают на достаточный пул прегенеративной группы и служат залогом стабильности популяции. С другой стороны, при расчёте Iвозраст не учитывается генеративная группа, а в случае завершения жизненного цикла в g3 возрастном состоянии (что также характерно и для C. purpurea) может давать некоторые искажения в интерпретации индекса. Индексы генеративности (Iг) для всех изученных популяций имеет достаточно высокие значения (Iг – 21,43 – 83,87).
Более детальная характеристика прослеживается при рассмотрении популяций в классификации «дельта-омега». В пространстве «дельта-омега» исследуемые популяции относятся к молодым (ЦП 2, 4, 5), переходным (ЦП 1, 3, 6), зреющим (ЦП 8, 11) и зрелым (ЦП 9, 10), (рис. 7). Отсутствие стареющих и старых групп популяций связано с коротким постгенеративным периодом C. purpurea и быстрым выпадением сенильных особей из популяции. В соответствии с классификацией Л. А. Жуковой, исследуемые ЦП относятся к нормальным, значения коэффициента возрастности находятся в пределах (Δ) – 0,23–0,51.
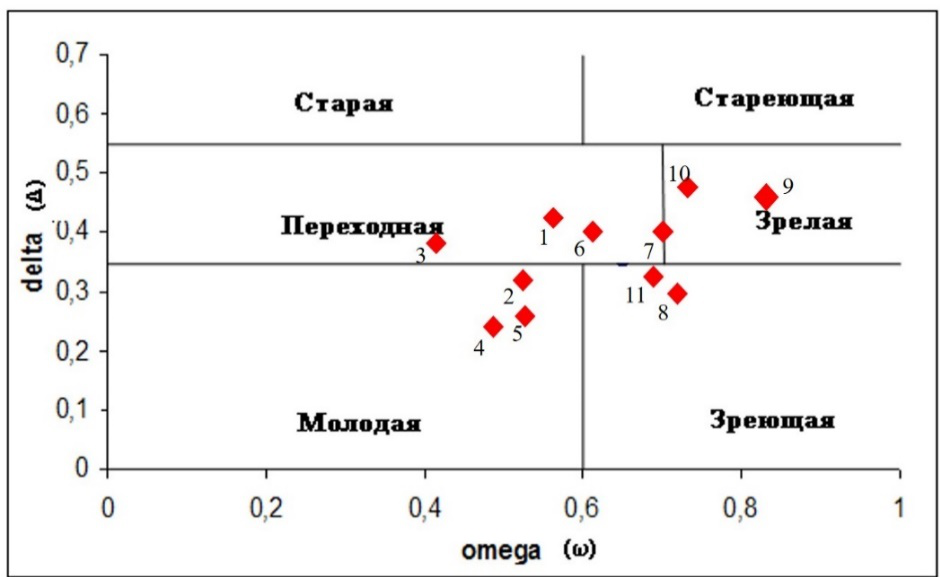
Рис. 7. Типы ценопопуляций Crepis purpurea по классификации «дельта-омега» (1–11– номера ценопопуляций).
Для исследования виталитетного анализа были выбраны наиболее вариабельные признаки – высота генеративного побега, длина и ширина листа.
Соотношение виталитетных классов показывает динамику увеличения участия «средних» и «высоких» классов (то есть от депрессивных к процветающим) при рассмотрении популяций от восточно-предгорных к западно-предгорным и далее – к яйлинским вариантам популяций (рис. 8). Так, к депрессивным относятся самая восточная (из обнаруженных) ЦП № 1, равновесными являются ЦП 2 и ЦП 3, произрастающие в восточном Предгорье (Белогорский район), а к процветающим принадлежат ЦП 4, 5, 7, 8, 10 и 11, обнаруженные в центральном Предгорье (Бахчисарайский район), на Чатырдаг-яйле и Гурзуфской яйле. Исключениями являются ЦП 6 (Бахчисарайский район) и ЦП 9 (Балаклавский район), которые находятся под дополнительной антропогенной нагрузкой (вблизи троп и пастбищ).
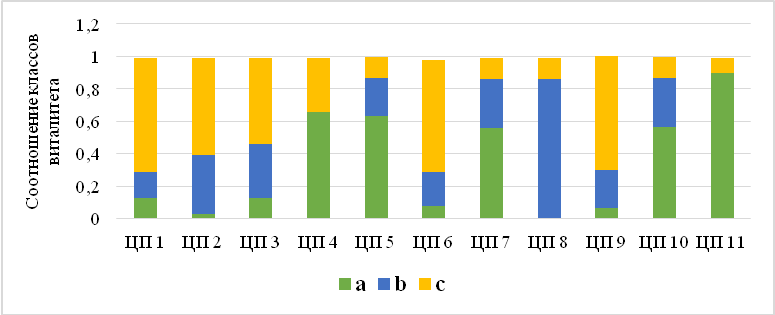
Рис. 8. Виталитетная структура ценопопуляций Crepis purpurea.
ЦП – ценопопуляция; а – высший класс; b – промежуточный класс; с – низший класс растений.
Наиболее заметна данная динамика при рассмотрении коэффициента качества популяции (Q). Повышение значений индекса Q, вероятно, зависит от повышения уровня осадков, связанных с изменением типов рельефа и высотности в различных точках западной и восточной частей Внутренней гряды, а также Главной гряды Крымских гор (рис. 9).
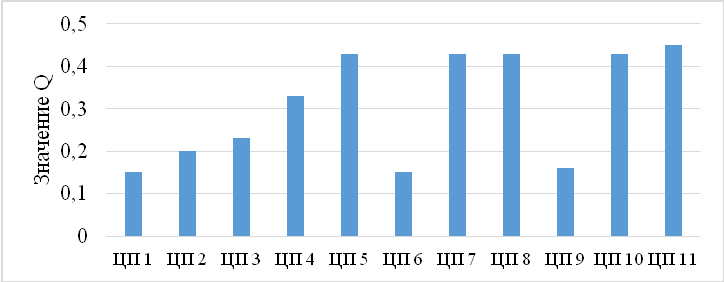
Рис. 9. Индекс виталитетного качества популяции Crepis purpurea.
Таким образом, в Предгорном Крыму C. purpurea ведет себя как ценофоб и развивает максимальные площади популяций на щебнисто-мергелистых склонах-подножьях с низким проективным покрытием травостоя, и приурочен к подножьям куэст и останцов Внутренней гряды Крымских гор. В этих условиях, как правило, формируются многочисленные (более 1000 особей), полночленные ЦП, с максимальной плотностью особей на 1 м2. Несмотря на благоприятные климатические условия произрастания на яйлах и южных склонах Главной гряды, о чем свидетельствуют высокие значения жизненности популяций, в этих условиях C. purpurea существует в виде небольших малоустойчивых популяций (низкая численность и плотность особей, неполночленность популяций) и, в силу ценофобности, легко вытесняется на более экстремальные экотопы – бровки скал, известняковые обрывы и очень крутые щебнисто-мергелистые склоны. К основным природным лимитирующим факторам развития популяций C. purpurea можно отнести эктопические (высокая подвижность склонов) и фитоценотические (высокое проективное покрытие травостоя).
Исследование пространственной структуры позволило выявить, что для большинства ценопопуляций C. purpurea характерен групповой тип распределения особей (значения IO ≥ 2), сохраняющийся при различных площадях популяций, численности особей и типах сообществ. Исключениями являются ЦП 7 (IO – 1,05) и ЦП 8 (IO – 1,86), (табл. 1).
Таблица 1
Характеристика пространственной структуры ценопопуляций Crepis purpurea
|
№ ЦП |
ПлощадьЦП, (м2) |
Проективное покрытие C. purpurea, (%) |
Численность особей C. purpurea |
Плотность C. purpurea на м2 |
Индекс Одума (IO) |
Характер пространствен-ной структуры |
|
1 |
1,180 |
3% |
Более 1000 особей |
7,10 |
2,38 |
групповое |
|
2 |
1,380 |
4% |
Более 1000 особей |
7,51 |
4,65 |
групповое |
|
3 |
588 |
5% |
Более 1000 |
8,03 |
2,41 |
групповое |
|
4 |
4,897 |
5% |
Более 1000 особей |
8,00 |
3,87 |
групповое |
|
5 |
470 |
3% |
210 |
2,06 |
2,26 |
групповое |
|
6 |
592 |
1% |
35 |
1,16 |
4,79 |
групповое |
|
7 |
143 |
2% |
68 |
1,20 |
1,05 |
случайное |
|
8 |
10 |
0,3% |
6 |
0,23 |
1,86 |
случайное |
|
9 |
10 |
0,5% |
27 |
0,95 |
4,45 |
групповое |
|
10 |
10 |
2% |
12 |
0,56 |
3,36 |
групповое |
|
11 |
5 |
0,5 |
9 |
0,33 |
3,17 |
групповое |
ЗАКЛЮЧЕНИЕ
Crepis purpurea по типу биоморфы – слабо партикулирующий стержнекорневой травянистый поликарпик с общей продолжительностью онтогенеза 5–6 лет (А–тип и подтип А 2), который проходит одна вегетативно неподвижная особь с осуществлением постгенеративного периода.
Для C. purpurea характерен структурный тип поливариантности с формированием 3 вариантов онтобиоморфы: особей с одно-, дву- или многоглавым каудексом.
Базовый онтогенетический спектр изученных популяций C. purpurea бимодальный с пиком на виргинильных (v) и зрелых генеративных особях (g3). В пространстве «дельта-омега» популяции относятся к молодым, переходным, зреющим и зрелым.
Особи C. purpurea тяготеют к групповому распределению в популяции, что подтверждается высокими значениями индекса Одума (2,26-4,65).
В виталитетных классах прослеживается динамика по увеличению «средних» и «высоких» классов от восточно-предгорных к западно-предгорным и далее – к яйлинским вариантам популяций.
REFERENCES
- Zlobin Y. A., Sklyar V. G., Klimenko A. A. Populations of rare plant species: theoretical foundations and research methods, 263 p. (Universitetska kniga,Sumy, 2013).
- Abdulganieva E. F., Vakhrusheva L. P. Phytocenotic confinement and distribution of Crepis purpurea (Willd.) M. Bieb. [Lagoseris purpurea (Willd.) Boiss.] in the Foothill Crimea. IV Scientific and Practical Conference of the Faculty, Graduate Students, Students and Young Scientists «Days of Science of the KFU named after V.I. Vernadsky». V. 2, P. 1248–1249, (2018).
- Abdulganieva E. F., Vakhrusheva L. P. Population Status Crepis purpurea (Willd.) M. Bieb. [Lagoseris purpurea (Willd.) Boiss.] On the southern border of the range (Turkey). V Scientific and Practical Conference of the Faculty, Graduate Students, Students and Young Scientists «Days of Science of the KFU named after V.I. Vernadsky». Botany and plant protection, P. 3–4, (2019).
- The Red Data Book of Sevastopol. Ed. by Dr. Sci., Prof. I. V. Dovgal, Dr. Sci., Prof. prof. V. V. Korzhenevsky. 432 p., (Publishing House ROST-DOAFK, Kaliningrad, Sevastopol, 2018).
- Red book of the Republic of Crimea. Plants, algae and fungi Ed. by Dr. Sci., Prof. Yena A. V. & Dr. Faterygа A. V., 480 p. (ARIAL LLC, Simferopol, 2015).
- Red Data Book of Turkish Plants (Pterdophyta and Spermatophyta), Edited by Ekim T., Koyuncu M., Vural M., Duman H., Aytac Z., Adiguzel N., 246 p. (Ankara, Türkiye Tabiatını Koruma Derneği, Van 100. yıl Üniversitesi, 2000).
- Convention on the Conservation of European Wildlife and Natural Habitats, Appendix I – Strictly protected flora species / URL: https://rm.coe.int/1680078aff (date of the application: 20.04.2020).
- Rabotnov T. A. The Life Cycle of Perennial Herbaceous Plants in Meadow Cenoses, Works of BIN AN SSSR, 3, 6, 7. (1950).
- Uranov A. A. Age range of coenopopulations as a function of time and energy wave processes, Biological Sciences, 2, 7 (1975).
- Smirnova O. V., Zagolgova L. B., Ermakova I. M., Plant populations (basic concepts and structure), 216 p. (Nauka, Moscow, 1976).
- Zlobin Yu. A. Principles and methods of studying coenotic plant populations, 146 p (KGU Publishing House Kazan, 1989).
- Zhukova L. A., Glotov N. V. The diversity of ontogenesis pathways in plant populations, Ontogenesis, 6, 455 (2001).
- Zhukova L. A. Ontogenesis and reproduction cycles of plants. Journal of General Biology, 44, (3), 361 (1983).
- Zhukova L. A. Population life of meadow plants, 224 p. (RIIK «Lanar», Yoshkar-Ola, 1995).
- Zhukova L. A., Notov A. A. On the role of the population-ontogenetic approach in the development of modern biology and ecology, Bulletin of Tver State University. Biology and ecology, 32, (31), 293 (2013).
- Kovalenko I. M. The structure of the population, which dominant ground layer of woody of phytocoenosis at National natural park Desnyansko-Starogutsky. I. Ontogenetic structure, Ukrainian Botanical Journal, 62, 5, 707 (2005).
- Zhivotovsky L. A. Ontogenetic states, effective density and classification of plant populations, Ecology, 21 (2001).
- Zlobin Yu. A. On the inequality of individuals in plant populations, Botanical Journal, 65, 3, 311 (1980).
- Odum Yu., Ecology, 1, 328 p. (Mir Publishing House, Moscow, 1986).
- Yena A. V. Spontaneous flora of the Crimean Peninsula, 232 p. (N. Orianda, Simferopol, 2012).
- Golubev V. N. The biological flora of the Crimea, 126 p. (GNBS, Yalta, 1996).
- Cherepanov S. K., Crepis L., Lagoseris M. B. Flora of the USSR (Ed. by Bobrov E. G., Tsvelev N. N), 29, 271 p. (Nauka, Moscow, Leningrad, 1964).
- Wulf E. V. Flora of the Crimea (Ed. by N. I. Rubtsov, L. A. Privalova), 3, (3), 393 p. (Yalta: Publishing House of the GNSS, 1969).
- Nikiforov A. R. Aging population repertoire features of Lagoseris callicephala Juz. (Asteraceae), Bull. of the State Nikita Botan. Gard., 125, 83 (2017).
- Nikiforov A. R., Nikiforova A. A. Rhythmological differences in development of Lagoseris callicephala and Lagoseris purpurea (Asteraceae), Bull. of the State Nikit. Botan. Gard., 118, 58 (2016).
- Lesina S. A., Koroteeva E. V. Ontogenesis and growing ecology of Scorzonera glabra in the Chelyabinsk region, Bulletin of the Orenburg State University, 12, 202 (2011).
- Modern approaches to the description of the plant structure Ed. N. P. Savinykh and Yu. A. Bobrov, 275 (Kirov, 2008).
- Pichugina E. V. Biomorphology and structure of coenopopulations of Jurinea cyanoides (L.) Reichenb. and Dianthus arenarius L. in the north-east of European Russia in connection with their protection, 261 p. (Kirov, 2007).
- Zaugolnova L. B., Denisova L. V., Nikitina S. V. Approaches and assessment of the state of plant populations, Bulletin of the Moscow Society of Naturalists. Department of Biology, 98, 5, 100 (1993).
