
CYTOGENETIC MARKERS FOR SELECTION MATERNAL TREES OF SCOTS PINE AND WHITE SPRUCE PRODUCING SEED OFFSPRING WITH CERTAIN LEVEL OF GENETIC STABILITY
JOURNAL: «SCIENTIFIC NOTES OF V.I. VERNADSKY CRIMEAN FEDERAL UNIVERSITY. Biology. Chemistry» Volume 7 (73), №3, 2021
Publication text (PDF):Download
UDK: 575.174.015.3:582.475.4/.2
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Kalaev V. N., Voronezh State University, Voronezh, Russian Federation
Ignatova I. V., Voronezh State University, Voronezh, Russian Federation
Kharchenko N. N., Voronezh State Forestry University named after G.F. Morozov, Voronezh, Russian Federation
Karpova S. S., Voronezh State Institute of Physical Culture, Voronezh, Russian Federation
TYPE:Article
DOI:https://doi.org/10.37279/2413-1725-2021-7-3-55-67
PAGES:from 55 to 67
STATUS:Published
LANGUAGE:Russian
KEYWORDS: Scots pine, white spruce, cytogenetic predictor, stability of genetic material, cytogenetic parameters.
ABSTRACT (ENGLISH):
Currently, the depletion of the gene pool of forest woody plants is observed due to the fact that in the course of selective felling, specimens valuable for economic characteristics were destroyed and seeds from the remaining specimens were used, which did not have the best qualities. It was noted that a high level of genetic heterogeneity allows the population to adapt to changing environmental conditions, ensures the stability of the population and is a resource for breeding. The selection of valuable genotypes in the creation of forest seed plantations, which is carried out according to the phenotype, is based on the heterogeneity of populations. Meanwhile, studies show that phenotypically normal trees do not always produce good offspring. The cytogenetic method allows assessing the offspring. With its help, it is possible to distinguish among the seed offspring the mother trees of the group with different levels of stability of the genetic material.
The aim of the work was to identify markers among cytogenetic parameters for the selection of trees producing seed offspring with different stability of the genetic material.
The objects of the study were seedlings of seeds of phenotypically normal Scots pine trees growing in the Khopersky State Nature Reserve (51°10’56.9″N 41°44’17.2″E), and white spruce growing in the Botanical Garden. prof. B.M. Kozo-Polyansky Voronezh State University (51°42’41.57 «N 39°12’17.57″E). The trees had no visible pest damage. The method of preparation and analysis of preparations of seed sprouts is described in the work of Butorina A.K. (2000). In the course of the study, 20 cytogenetic parameters were determined: the mitotic index (counted with and without taking into account cells at the prophase of mitosis), the level of mitotic pathologies (counted with and without taking into account cells at the prophase of mitotic), the proportions of cells at the stages of prophase, metaphase, anaphase , the proportions of cells with 1–10 nucleoli in the nucleus of interphase cells, the proportion of cells with micronuclei, the proportion of cells with residual nucleoli in interphase and mitosis. Statistical processing of the study results was carried out using the STADIA 7.0 and MedCalc 17.5.3 software. The diagnostic value of the indicator is characterized by the area under the ROC-curve: 0.9–1.0 – excellent; 0.8–0.9 – very good; 0.7–0.8 – good, 0.6–0.7 – average, 0.6 and less – unsatisfactory.
Among the cytogenetic indicators, there are those that are suitable for separating the mutable group from the weakly mutable and intermediate, as well as the weakly mutable from the intermediate. In pine, these include the proportion of cells at the prophase stage and the average number of nucleoli in the nucleus, in spruce – the mitotic index without taking into account cells at the prophase stage, the level of mitotic pathologies with and without taking into account cells at the prophase stage and the proportion of cells at the metaphase stage. There are indicators that make it possible to distinguish the mutable group from the weakly mutable and from the intermediate, but are not suitable for separating the weakly mutable group from the intermediate. These include the mitotic index, calculated taking into account cells at the prophase stage in pine, and the proportion of cells at the metaphase and ana-telophase stages in spruce. According to some indicators, it is possible to distinguish the intermediate group from the mutable and the weakly mutable, but it is impossible to distinguish the mutable group from the weakly mutable. In pine, such indicators are the proportion of cells in the metaphase, and in spruce, the proportion of cells with residual nucleoli in the interphase and meta-telophase. Indicators have been identified that make it possible to distinguish only mutable group from weakly mutable (in pine, the proportion of cells at the ana-telophase stage), and weakly mutable group from intermediate (in pine, the mitotic index taking into account cells at the prophase stage, the level of mitotic pathologies with and without taking into account cells at prophase stage, in spruce – mitotic index, taking into account cells at the prophase stage, the proportion of cells with micronuclei).
ВВЕДЕНИЕ
В связи с меняющимся климатом на планете остро встает вопрос сохранения генетических ресурсов растений. В настоящее время наблюдается обеднение генофонда лесных древесных растений из-за то, что в ходе выборочных рубок шло уничтожение ценных по хозяйственным признакам экземпляров и использование семян от оставшихся экземпляров, которые не обладали лучшими качествами. К обеднению генофонда приводит также использование семян из других районов. Было замечено, что высокий уровень генетической гетерогенности позволяет популяции адаптироваться к изменяющимся условиях окружающей среды, обеспечивает стабильность популяции [1, 2] и является ресурсом для проведения селекционных работ [3]. На гетерогенности популяций основывается отбор ценных генотипов при создании лесосеменных плантаций, который проводят по фенотипу [4]. Между тем исследования показывают, что от фенотипически нормальных деревьев не всегда получается хорошее потомство. Провести оценку потомства позволяет цитогенетический метод [5]. С его помощью можно выделить среди семенного потомства материнские деревья с разным уровнем стабильности генетического материала. Подобные работы выполнены в Воронежской области по дубу черешчатому, березе повислой, сосне обыкновенной, ели белой, рододедрону Ледебура, ели колючей, ореху грецкому [6–14]. Во всех работах даны характеристики выделенных групп, но отсутствует формальное обоснование выбора маркеров, которые позволили бы отнести проросток к той или иной группе. Для решения данной задачи используют методы ROC-анализа, который позволяет среди множества исследуемых характеристик выбрать предикторы (показатель, который позволяет разделить выборку на группы с заданными характеристиками), позволяющие отнести каждый конкретный объект к той или иной группе [15].
Целью работы стало выявление среди цитогенетических показателей маркеров (предикторов) для отбора деревьев, продуцирующих семенное потомство с разной стабильностью генетического материала.
МАТЕРИАЛЫ И МЕТОДЫ
Объектами исследования явились проростки семян фенотипически нормальных деревьев сосны обыкновенной, произрастающих в Хоперском государственном природном заповеднике (51°10’56.9″N 41°44’17.2″E), и ели белой, произрастающих в ботаническом саду им. проф. Б.М. Козо-Полянского Воронежского государственного университета (51°42’41.57″N 39°12’17.57″E). Деревья не имели видимых повреждений вредителями. Методика приготовления и анализа препаратов проростков семян изложена в работе [16]. В ходе исследования определяли 20 цитогенетических показателей: митотический индекс (с учетом и без учета клеток на стадии профазы митоза), уровень патологий митоза (с учетом и без учета клеток на стадии профазы митоза), доли клеток на стадиях профазы, метафазы, анафазы, доли клеток с 1–10 ядрышками в ядре интерфазных клеток, долю клеток с микроядрами, долю клеток с остаточными ядрышками в интерфазе и митозе. Статистическую обработку результатов исследования проводили при помощи программ STADIA 7.0 и MedCalc 17.5.3 [15, 17]. Диагностическую ценность предиктора характеризует площадь под ROC-кривой: 0.9–1.0 – отличная; 0.8–0.9 – очень хорошая; 0.7–0.8 – хорошая,
0.6–0.7 – средняя, 0.6 и меньше – неудовлетворительная.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Ранее методами кластерного анализа у исследуемых видов выделили проростки с разным уровнем стабильности генетического материала [7, 9]. Рядом авторов [8, 12–14] было показано, что проростки с низким уровнем нарушений митоза растут быстрее и имеют лучшие морфологические показатели по сравнению с проростками с высоким уровнем патологий митоза.
Для выявления цитогенетических предикторов генетической стабильности был использован ROC-анализ. Данный метод позволяет отделить одну группу от другой, но не всегда дает возможность провести границы между всеми исследуемыми группами. Такие показатели, как доля клеток с 1–10 ядрышками в ядре, не позволяют разделить проростки на группы по степени стабильности генетического материала у обоих исследуемых видов.
ROC-кривые предикторов, позволяющие отделить проростки мутабильной группы от слабомутабильной, представлены на рис. 1 и 2. Как видно из рисунков, у ели количество цитогенетических показателей, по которым можно разделить две группы проростков, больше, чем у сосны. Такие показатели, как доля клеток на стадии профазы и ана-телофазы митоза позволяют провести разделение групп у двух исследуемых видов растений.
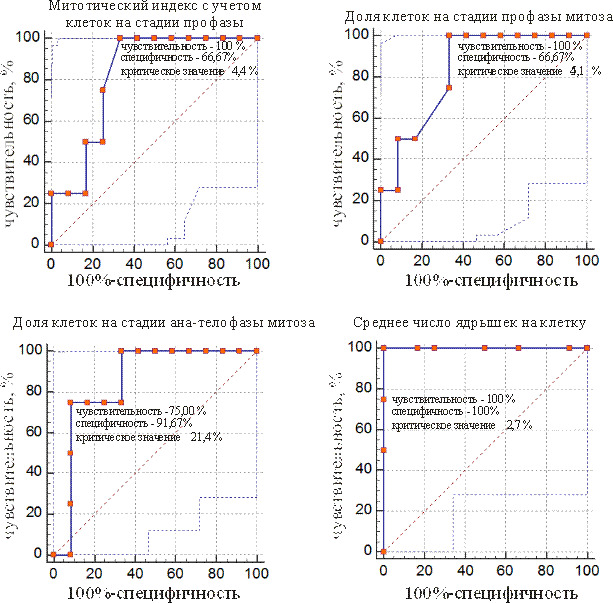
Рис. 1. ROC-кривые предикторов для разделения мутабильной и слабомутабильной групп проростков у сосны обыкновенной
У сосны отличным диагностическим критерием мутабильной группы является среднее число ядрышек на клетку, остальные цитогенетические признаки показали себя как очень хорошие диагностические критерии.
У ели белой хорошими показателями выступают митотический индекс, подсчитанный с учетом клеток на стадии профазы митоза, доля клеток на стадии метафазы и ана-телофазы митоза; остальные цитогенетические критерии являются очень хорошими диагностическими показателями.
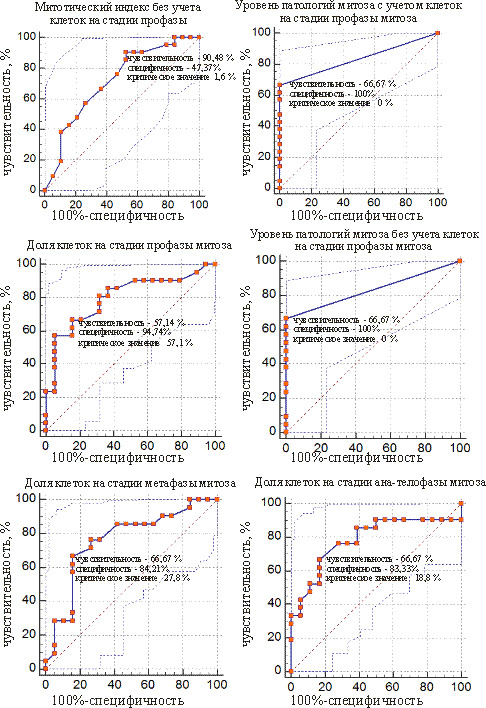
Рис. 2. ROC-кривые предикторов для разделения мутабильной и слабомутабильной групп проростков у ели белой
ROC-кривые предикторов, позволяющие отделить слабомутабильную группу от промежуточной, представлены на рис. 3 и 4. У сосны обыкновенной таких показателей 6, а у ели белой – 8. Такие показатели, как митотический индекс, подсчитанный без учета клеток на стадии профазы, уровень патологий митоза, подсчитанный с учетом и без учета клеток на стадии профазы, доля клеток на стадии метафазы, у обоих видов могут служить для отделения проростков из слабомутабильной группы от проростков из промежуточной группы.
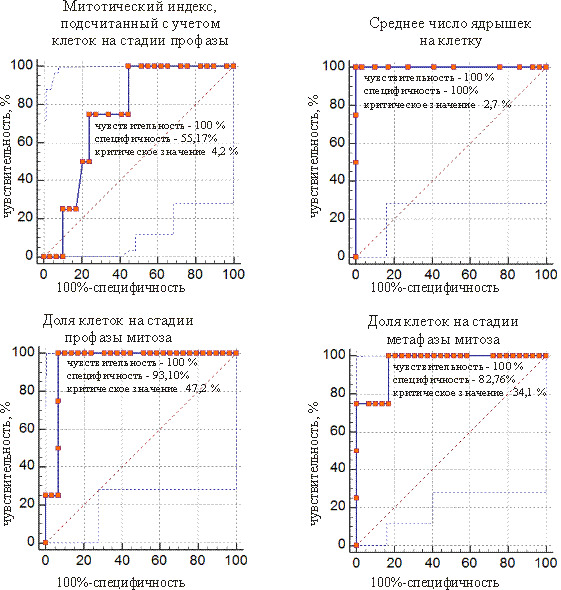
Рис. 3. ROC-кривые предикторов для разделения мутабильной и промежуточной групп проростков у сосны обыкновенной
ROC-кривые предикторов для отделения проростков из мутабильной группы от проростков из промежуточной группы представлены на рис. 5 и 6. У сосны это 4 показателя, у ели – 8. Общими для двух видов являются предикторы «доля клеток на стадии профазы» и «доля клеток на стадии метафазы».
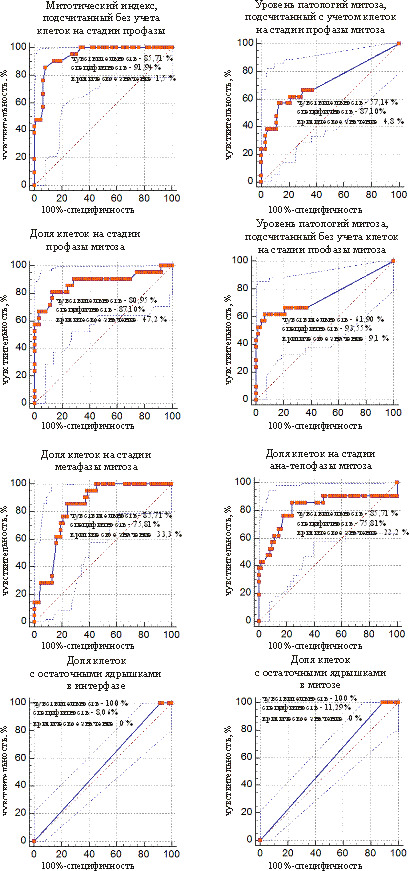
Рис. 4. ROC-кривые предикторов для разделения мутабильной и промежуточной групп проростков у ели белой
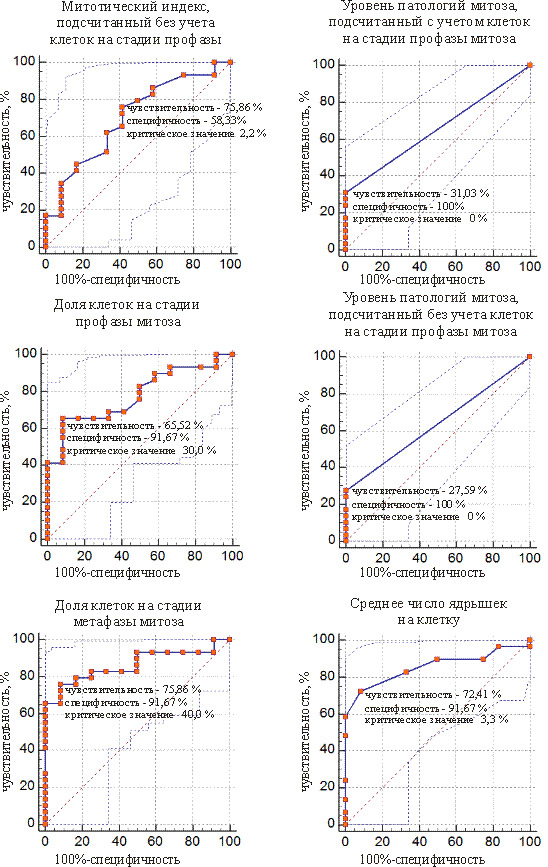
Рис. 5. ROC-кривые предикторов для разделения слабомутабильной и промежуточной групп проростков у сосны обыкновенной
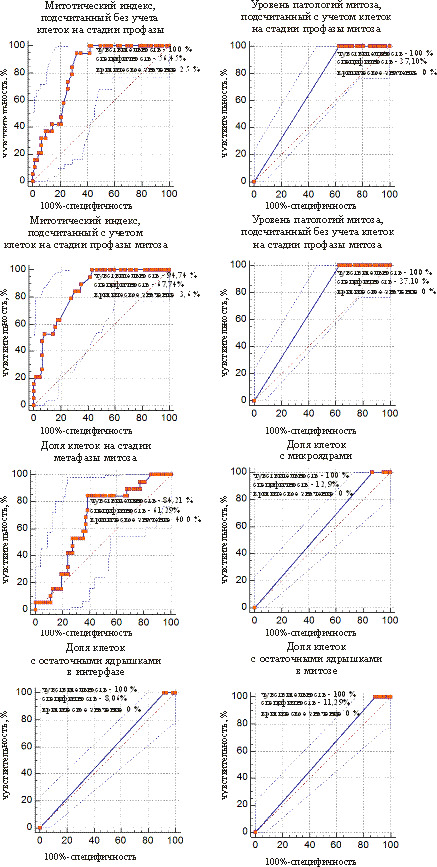
Рис. 6. ROC-кривые предикторов для разделения слабомутабильной и промежуточной групп проростков у ели белой
Среди цитогенетических показателей есть такие, которые подходят для отделения мутабильной группы от слабомутабильной и промежуточной, а также слабомутабильной от промежуточной. У сосны к ним относятся доля клеток на стадии профазы и среднее число ядрышек в ядре, у ели – митотический индекс без учета клеток на стадии профазы, уровень патологий митоза с учетом и без учета клеток на стадии профазы и доля клеток на стадии метафазы. Есть показатели, которые позволяют отграничить мутабильную группу от слабомутабильной и от промежуточной, но не подходят для разделения слабомутабильной и промежуточной групп. К их числу у сосны относятся митотический индекс, подсчитанный с учетом клеток на стадии профазы, у ели – доля клеток на стадии метафазы и ана-телофазы. По некоторым показателям можно отграничить промежуточную группу от мутабильной и слабомутабильной, но нельзя отграничить мутабильную группу от слабомутабильной. У сосны такими показателями выступают доля клеток в метафазе, а у ели – доля клеток с остаточными ядрышками в интерфазе и мета-телофазе. Выявлены показатели, которые позволяют отграничить только мутабильную от слабомутабильной (у сосны – доля клеток на стадии ана-телофазы), и слабомутабильную от промежуточной (у сосны – митотический индекс с учетом клеток на стадии профазы, уровень патологий митоза с учетом и без учета клеток на стадии профазы, у ели – митотический индекс с учетом клеток на стадии профазы, доля клеток с микроядрами).
ЗАКЛЮЧЕНИЕ
На основании выполненных исследований можно рекомендовать проводить отбор деревьев, продуцирующих семенное потомство с разным уровнем стабильности генетического аппарата, по цитогенетическим показателям, представленным в таблице.
Для того, чтобы дать оценку семенного потомства дерева, необходимо определить цитогенетические показатели проростков семян (минимальное количество 10 штук). Каждый исследуемый проросток относят к одной из трех групп (по большинству показателей). Затем считают количество проростков в каждой группе (слабомутабильной, мутабильной и промежуточной) и дают рекомендации о дальнейшем использовании семенного потомства изучаемого дерева.
Таблица
Значения цитогенетических показателей, позволяющих отнести проросток к группе с разным уровнем стабильности генетического материала
| Цитогенетический показатель | Сосна обыкновенная | Ель белая | ||||
| слабо-мутабиль-ная | промежу-точная | мутаби-льная | слабо-мутабиль-ная | промежу-точная | мутаби-льная | |
| Митотический индекс, подсчитанный с учетом клеток на стадии профазы митоза,% | <4,4 | <4,2 | >4,4
>4,2 |
≤3,6 | ≥3,6 | – |
| Митотический индекс, подсчитанный без учета клеток на стадии профазы митоза, % | >2,2 | <2,2 | 1,6–2,5 | ≥2,5 | <1,6 | |
| Уровень патологий митоза, подсчитанный с учетом клеток на стадии профазы митоза, % | 0 | >0 | – | 0 | <4,8 | >4,8 |
| Уровень патологий митоза, подсчитанный без учета клеток на стадии профазы митоза, % | 0 | >0 | – | 0 | ≤9,1 | ≥9,1 |
| Доля клеток на стадии профазы, % | 30–46,1 | ≤47,2
≤30 |
>47,2
>46,1 |
<57,1 | <47,2 | >57,1
>47,2 |
| Доля клеток на стадии метафазы, % | >40 | 34,1–40 | <34,1 | 27,8–40 | >40 | <27,8 |
| Доля клеток на стадии ана-телофазы, % | >21,4 | – | <21,4 | >18,8 | >22,2 | <18,8
<22,2 |
| Среднее число ядрышек в ядре, шт. | 2,7–3,3 | >2,7
>3,3 |
<2,7 | – | – | – |
REFERENCES
- MilyutinI. On some problems of preserving the forest genetic resources of Siberia. Abstracts of the III Intern. meetings «Conservation of genetic resources of Siberian forests» (Krasnoyarsk: KrasGAU, 2011), p. 94.
- Booy, Hendriks R. J. J., Smulders M. J. M., Van Groenendael J. M., Vosman B. Genetic diversity and the survival of populations, Plant biology, 2, 379 (2000).
- Ivetić , Devetaković J. Concerns and evidence on genetic diversity in planted forests, Reforesta, 3, 196 (2017).
- Selection of plus trees and plantations: methodical. instructions (comp. N.V. Laur, A.P. Tsarev., Petrozavodsk, 2005), p. 36.
- VostrikovaV. The area of using the cytogenetic method in forest genetics and breeding on the example of silver birch, Forestry, 1, 30 ( 2006).
- BurmenkoV., Baranova T. V., Kalaev V. N., Sorokopudov V. N. Cytogenetic polymorphism of seed progeny of introduced plants on the example of Rhododendron ledebourii Pojark., Turczaninowia, 21 (1), 164 (2018).
- KalaevN., Artyukhov V. G., Popov V. N., Ignatova I. V. Cytogenetic polymorphism of pine seeds in eastern Voronezh region, Russian Forest Sciences, 4, 56 (2010).
- KalaevN., Popova A. A. Cytogenetic characteristics and morphological parameters of english oak seed progeny oak (Quercus robur L.), growing in territories with different levels of anthropogenic pollution, Bulletin of the Voronezh State University. Series: Chemistry. Biology. Pharmacy, 4, 63 (2014).
- KalaevN., Ignatova I. V., Kalaeva E. A. Cytogenetic polymorphism of seed progeny of white spruce trees (Picea glauca (Moench) Voss) during the introduction in the Voronezh region, Turczaninowia, 23 (3), 185 (2020).
- KarpovaS. Influence of anthropogenic pollution on cytogenetic polymorphism of seed progeny of silver birch (Betula pendula Roth) in natural and artificial stands: dis. … cand. biol. sciences. (Voronezh, 2011), p. 149.
- KarpovaS., Al-Hachami F. R. H., Kalaev V. N., Ignatova I. V., Slavsky V. A. Cytogenetic polymorphism of walnut trees (Juglans regia L.) seed progeny during introduction in the Voronezh region, Ecology of urbanized territories, 2, 21 (2020).
- BaranovaV., Zemlyanukhina O. A., Kalaev V. N. Pat. No. 2716112 Method for selection of Betula pendula maternal trees producing seed offspring with different germination and stability of the genetic material of somatic cells, according to biochemical parameters, No. 2019115745; declared 05/22/2019; publ. 05.03.2020, Bul. No. 7. p. 15.
- BaranovaV., Kalaev V. N. Pat. 2681105 Russian Federation, IPC A01H 1/04 (2006.01) Method for selection of mother plants Picea pungens Engelm. Producing seed offspring with different levels of stability of genetic material and better morphometric parameters, No. 2018114226; declared 04/17/2018; publ. 04.03.2019, Bul. No. 7. p. 17.
- BaranovaV., Kalaev V. N., Burmenko Yu. V. Pat. 2654605 Russian Federation, IPC A01H 1/04 (2006.01) A01H 5/00 (2006.01) Method for assessing the quality of seeds of Rhododendron Ledebourii Pojark by cytogenetic parameters, No. 2016140096; declared 10/11/2016; publ. 05/21/2018, Bul. No. 15. p. 16.
- DeLongR. DeLong D. M., Clarke-Pearson D. L. Comparing the areas under two or more correlated receiver operating characteristic curves: a nonparametric approach, Biometrics, 44, 837 (1988).
- ButorinaK., Kalaev V. N. Analysis of sensitivity of different criteria in cytogenetic monitoring, Russian Journal of Ecology, 31 (3), 186 (2000).
- KalaevaA. Artyukhov V. G., Kalaev V. N. Theoretical foundations and practical application of mathematical statistics in biological research and education: textbook (Voronezh: Voronezh State University Publishing House, 2016), p. 282.
