
MORPHOLOGICAL CRITERIA FOR THE DIFFERENTIATION OF AGE STATUS OF PTILOSTEMON ECHINOCEPHALUS (WILLD.) GREUTER (LAMYRA ECHINOCEPHALA (WILLD.) TAMAMSCH.)
JOURNAL: «SCIENTIFIC NOTES OF V.I. VERNADSKY CRIMEAN FEDERAL UNIVERSITY. Biology. Chemistry» Volume 7 (73), №2, 2021
Publication text (PDF): Download
UDK:598.1:591.53 (477.75)
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Zadneprovskaya E. V., Timiryazev Institute of Plant Physiology of RAS, Moscow, Russia
Vakhrusheva L. P., V. I. Vernadsky Crimean Federal University, Simferopol, Crimea, Russian Federation
TYPE: Article
DOI: https://doi.org/10.37279/2413-1725-2021-7-2-58-68
PAGES: from 58 to 68
STATUS: Published
LANGUAGE: Russian
KEYWORDS:Ptilostemon echinocephalus, morphological criteria, age states, phyyganoid phytocoenoses, petrophytic steppes, Crimea
ABSTRACT (ENGLISH):
Ptilostemon echinocephalus (Willd.) Greuter (Lamyra echinocephala (Willd.) Tamamsch.) is the only species of the genus Ptilostemon from the Asteraceae family that grows on the Crimean Peninsula.
The relic area of Ptilostemon echinocephalus distributed by the Crimea, the Caucasus, Asia Minor. The species is included in the Red Book of the Republic of Crimea, Krasnodarskiy and Stavropolskiy Krays of the Russian Federation. The article is the result of a long-term study of morphological criteria of Ptilostemon echinocephalus age status. Thus, the ontogenesis of individuals of the species Ptilostemon echinocephalus includes 12 age states, the total during of ontogenesis lasts 15–18 years.
The main characters of the pregenerative period differentiation of development are the number of leaves and the differentiation degree of the leaves lamina. During the study, another important feature was identified to indicate different age stages, the ratio of vegetating and dried leaves.
Generative plants were subdivided into: immature generative (g1), adult (mature) generative (g2) and old generative (g3). For the generative stage of development, the ratio of generative and vegetative rosettes, the ratio of dried and vegetative shoots, and the number of baskets were indicative.
Differentiation of subsenile and senile age states is conducted according to the same criteria that are used for generative individuals, the main emphasis is on the ratio of dried and vegetative parts.The article also notes the specificity of the course of ontogenesis depending on the ecological-cоenotic conditions in the petrophytic steppes and phrygana communities of the Crimea.
ВВЕДЕНИЕ
Ptilostemon Cass. (incl. Lamyra (Cass.) Cass., Chamaepeuce DC. – род семейства Asteraceae, включающий, согласно данным Международного альянса Asteraceae (TICA) [1] и международной базе данных The Plant List [2], 18 видов растений, произрастающих на Крымском полуострове, Кавказе, в Южной Европе и Малой Азии [3].
На территории Крыма и Российской Федерации обнаружен только один вид данного рода – Ptilostemon echinocephalus (Willd.) Greuter (Lamyra echinocephala (Willd.) Tamamsch.) – реликтовый вид. Ареал Ptilostemon echinocephalus охватывает Крым, Кавказ, Малую Азию. Он внесен в Красную книгу Республики Крым [4], а также в Красные книги Краснодарского [5] и Ставропольского краев [6, 7].
Ptilostemon echinocephalus показывает достаточно широкую фитоценотическую амплитуду в пределах Горного Крыма, встречаясь от петрофитных степей до фриганоидных фитоценозов, о чем свидетельствуют публикации В. В. Корженевского и Л. Э. Рыфф [8, 9], а также наши предшествующие исследования [10, 11], позволившие первично установить некоторые онтогенетические признаки вида и его фитоценотическую приуроченность. Однако, несмотря на указанный в ККРК статус «2» (редкий) [4], с популяционной точки зрения Ptilostemon echinocephalus остается малоизученным. Адаптивная стратегия вида и возможность успешно существовать в разнообразных эколого-ценотических условиях, как известно, проявляется через признаки популяционной структуры и, прежде всего, посредством особенностей онтогенетической структуры. Предварительной ступенью для ее изучения является нахождение комплекса морфологических критериев, характеризующих различные онтогенетические состояния, что и составляет цель настоящего исследования.
МАТЕРИАЛ И МЕТОДЫ
Основная часть исследований проводилась в фитоценозе петрофитной степи Stipeto-Satureetum festucoso-lamiriosum на территории ландшафтно-рекреационного парка «Битак», в окр. Симферополя [10, 11] а также в петрофитных степях и фриганоидных сообществах Долгоруковской яйлы, Чатырдага, Мангуп Кале в 2017–2020 гг.
Выявление и учет качественно-количественных критериев морфологических признаков возрастных состояний особей Ptilostemon echinocephalus осуществлялись в соответствии с классическими методами [12–16].
Проростки и ювенильные особи наблюдали как в природных условиях, так и в условиях искусственного выращивания. Проращивание семян осуществлялось согласно ГОСТу [17], однако в связи с малым количеством семян и редкостью растений, число заложенных для опыта семянок на чашку Петри ограничивалось 10–25 шт. Проклюнувшиеся семена высаживали в смесь торфа, агроперлита и вермикулита (1:1:1).
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Ранее возрастные состояния P. echinocephalus (L. echinocephala) были предварительно подразделены на периоды онтогенеза: латентный, прегенеративный, генеративный и сенильный [11]. В настоящей статье представлены разработанные авторами схематические изображения прегенеративной фазы развития P. echinocephalus, с детализацией морфогенеза листа, а также уточнен комплекс качественно-количественных морфологических признаков для дифференциации растений прегенеративного и генеративного онтогенетических состояний.
Главными признаками дифференциации прегенеративного периода развития являются количество листьев и степень дифференциации листовой пластинки. Онтогенетические состояния в прегенеративном периоде имеют следующий вид.
Латентный период. Семена (se) гладкие, черные, овальные 3–5 мм длиной.
Прегенеративный период. Проростки (p) при первоначальном исследовании в естественных популяциях обнаружены не были, поэтому их изучение осуществлялось в лабораторных условиях, позднее – были найдены во фриганоидных фитоценозах (2020 г.). Проростки появлялись в течение 5–10 дней после посева, гипокотиль достигал длины до 3 см, развивая два семядольных листа ярко-зеленого цвета округло-овальной формы: длиной до 1,5 см и шириной – 0,3–0,6 см. В начале ранне-весенней вегетации (апрель–май) проростки легко обнаруживались в естественных фитоценозах, причем наиболее обильны они были во фриганоидных сообществах.
Ювенильное состояние (j). Переход проростков в ювенильное состояние происходил через 21–30 дней. В этом возрасте сохраняются семядольные листья, но в начальный период ювенильного возраста они остаются зелеными, по мере взросления – становятся буро-зелеными, а в зрелом ювенильном – черно-бурыми, полностью высыхая к моменту перехода в имматурную фазу онтогенеза. У ювенильных особей развивается 4–7 зеленых ланцетных листьев с острой верхушкой, длиной от 0,7 см до 3,5 см. Во всех исследуемых образцах вторая пара листьев (внутреннего круга розетки) превышает длину первой в 3 раза (рис.1 а, b).
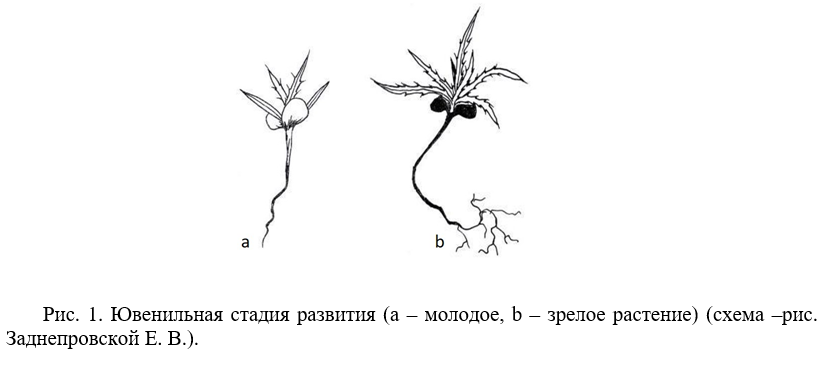
У молодых ювенильных растений лист ланцетовидной формы с ровным краем листовой пластинки и острой верхушкой. Затем на верхушке листа появляется «шипик», дальнейшая дифференциация происходит следующим образом: с обеих сторон листовой пластинки симметрично формируются «шипики», а к окончанию ювенильного возраста в месте локализации латеральных «шипиков» образуется зубчик листа (при этом острие сохраняется на каждом его зубчике) (рис. 2). В ювенильном возрастном состоянии растение пребывает до конца первого вегетационного сезона, а в конце переходит в молодое имматурное состояние и уходит в первую перезимовку. В случае влажного летнего сезона ювенильные растения продолжают формировать листья ювенильной биоморфы (до 15 шт.) и, в виде зрелого ювенильного растения, уходят в перезимовку. Таким образом, скорость прохождения фаз онтогенеза замедляется в условиях достаточного количества атмосферной влаги.

Общими закономерностями динамики развития молодых особей в составе растительных сообществ являются следующие: обычно проростки и ювенильные растения, появляясь обильно ранней весной, произрастают группами и поселяются рядом с материнскими растениями, но уже к имматурному возрасту происходит сильное изреживание и от группировок остаются единичные растения, которые в небольшом количестве сохраняются к началу–середине лета. В период сильной засухи, наступающей в августе, происходит еще одно отмирание молодых растений, что заметно сказывается на базовом онтогенетическом спектре, поскольку в нем всегда доминируют генеративные особи. Растения всех других возрастных состояний Ptilostemon echinocephalus оказываются малоустойчивыми для выживания в этих экстремальных условиях. В петрофитных степях проростки появляются также в заметном количестве ранней весной, локализуются на участках, лишенных растительного покрова или с малыми значениями проективного покрытия (10–12 %) других видов сообщества, поскольку в таких локусах снижена конкурентная напряженность. Однако в дальнейшем они массово элиминируются как вследствие усиливающейся с возрастом конкуренции, так и в связи с наступлением засушливого периода. В результате эти особенности находят выражение в различной пространственной структуре особей, формирующейся в петрофитных степях и во фриганодных сообществах.
Имматурное состояние (im). В этом возрасте у растения обнаруживаются от 7 до 14 зеленых вегетирующих листьев, длиной от 3,0 до 10,0 см, глубина рассечения листовых пластинок составляет 0,1–0,8 см. Чем более молодой лист, тем больше глубина рассечения его листа. Таким образом, зубчатый край листовой пластинки, сформировавшийся в ювенильном возрасте, преобразуется у имматурного растения в лопастной лист, каждая узкая лопасть которого заканчивается шипиком. В зависимости от особенностей прохождения ювенильной фазы, у имматурных особей сохраняются 8–15 сухих ювенильных листьев.
Найденные морфометрические отличия позволили дифференцировать особи на молодые имматурные (im1) и зрелые имматурные (im2) (табл. 1). Принципиальное отличие их морфологии в том, что у имматурной зрелой особи (в переходном возрастном состоянии к виргинильной) в области каудекса уже может иметь место начальная партикуляция, и тогда формируются особи с двумя розетками (рис. 4).
Таблица 1.
Морфометрические признаки имматурных растений P. echinocephalus
| Стадия | Количество вегетирующих листьев, шт. | Количество сухих листьев, шт. | Длина листа, см | Глубина рассечения листа, см | Длина корневой шейки |
| im1 | 7–12 | 8–12 | 3,0–7,5 | 0,1–0,5 | 1,5–4,7 |
| im2 | 10–14 | 10–15 | 6,0–10,0 | 0,4–0,8 | 5,0–8,5 |
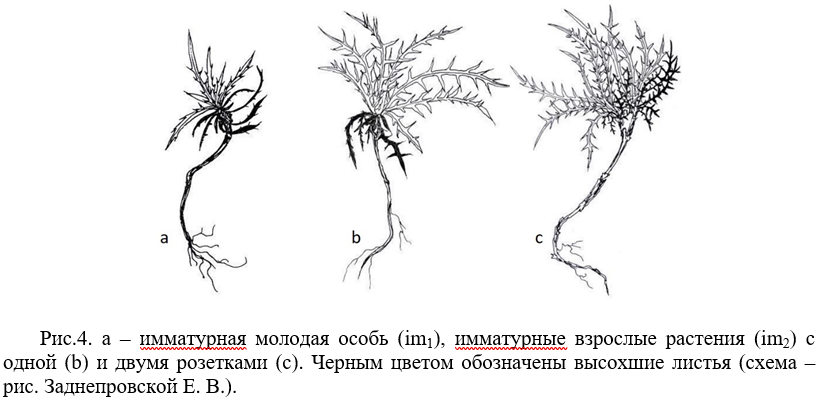
В имматурном возрасте P. echinocephalus может пребывать 1–2 года, уходя во вторую перезимовку как в молодом, так и во взрослом имматурном возрасте.
Виргинильное возрастное состояние (v). В следующую фазу онтогенеза Ptilostemon echinocephalus переходит на 2–3 год жизни и пребывает в ней до двух лет. В виргинильной фазе онтогенеза (рис. 5) количество отмерших листьев достигает 40–45, из них около 15 – это листья имматурного возраста, 25–30 засохших листьев, но уже с признаками глубины рассечения листовой пластинки, характерной для виргинильных растений. Вегетирующих зеленых листьев почти столько же, т.е. от 20 до 25 штук (длина зеленых листьев – 8–15 см), глубина рассечения листовой пластинки – 0,9–2,0 см. При таких особенностях морфологии листа, начиная с виргинильного возраста, его следует считать рассеченным (рис. 5). В случае, если виргинильное растение формирует две розетки, то процессы усыхания листьев идут еще интенсивнее, и засохших бывает в 2 раза больше, чем вегетирующих (Рис. 5b). Следовательно, для дифференциации возрастных состояний у P. echinocephalus, важно учитывать не только размер листьев и глубину (степень) рассечения листовой пластинки, но и такой признак как соотношение усохших и вегетирующих листьев, начиная с виргинильного возраста. Однако в классической биоморфологии рекомендуют данный прием использовать, начиная с генеративного и сенильного возрастов [13]. Вероятно, в связи с тем, что растения P.echinocephalus нередко обитают на чрезвычайно сухих экотопах – осыпных мергелистых склонах, часто и в условиях недостатка питательных веществ, где полностью отсутствует почвенный слой, а растения фактически растут на материнской породе, дефицит влаги и элементов питания «заставляет» их (ради выживания) вырабатывать особый, специфический механизм адаптации, который приводит к раннему усыханию листьев. Это обусловливает возникновение несоответствия у еще растущего растения в соотношении количества нарастающих и отмирающих частей растения.

Генеративный период. Генеративное возрастное состояние (g). Генеративный возраст у P.echinocephalus наступает на 4–5 год жизни. Генеративные растения были подразделены на: молодые генеративные (g1), взрослые (зрелые) генеративные (g2) и старые генеративные (g3). В нашей выборке молодые генеративные (g1) растения высотой до 30 см развивают от одной до трех розеток, образующих генеративные побеги с 1–6 корзинками (с фертильными цветками); вегетативных розеток 1–3 шт. Усохшие прошлогодние побеги и розетки с высохшими листьями присутствуют у растений этого возраста в числе 3–5 шт. Часть этих розеток может сохраниться от виргинильного возраста. Растение находится в молодом генеративном возрасте 2–3 года.
Взрослые (зрелые) генеративные (g2) показывают наибольшие показатели в развитии вегетативной и генеративной сфер. Растения g2 морфологически хорошо развиты, достигают высоты 30–50 см, развивают до 12 генеративных побегов, диаметр «куста» достигает 50–60 см. Максимальное количество корзинок на одном побеге – 6, все с фертильными цветками. Вегетативных розеток – 3 шт., розетки с высохшими листьями присутствуют в количестве 6 до 9 шт. (иногда до 10). Растение пребывает в этом возрасте до 4–5 лет. Здесь еще имеет место некоторое преобладание процессов роста над процессами отмирания (рис. 6b), в сообществах петрофитных степей примерно в соотношении 60/40. Зрелые генеративные растения доминируют также в исследованных фриганоидных сообществах. На склоновых экотопах у них развивается морфологическая адаптация в форме удлиненных ксилоризомов, которые обеспечивают растениям более надежное закрепление на динамичном грунте, слагаемом мергелистыми и мергелисто-глинистыми частицами эдафотопа. Кроме того, во фриганоидных сообществах у генеративных растений наблюдается соотношение сухих и зеленых листьев как 70/30, т.е. такое же, как это имеет место только у субсенильных и сенильных растений в петрофитных степях.

Что касается особей старого генеративного возраста (g3), то признаки, избранные нами для дифференциации, остаются теми же, что и для предыдущих возрастных состояний: высота растений, как правило, не превышает 30 см, формируется обычно не более двух генеративных побегов, каждый из которых несет 2–3 корзинки. Количество розеток с усохшими листьями достигает 10–12 шт.: к этому возрасту от многих из них остаются высохшие основания побегов с небольшим числом также высохших и частично разрушившихся листьев. Общий габитус растения начинает сильно меняться: происходит переход в состояние «рыхлого куста», т.е. вертикально стоящие побеги несколько отодвигаются друг от друга и приобретают наклонное положение в направлении от центра к периферии, в их прикорневой части формируется плагиотропная часть. Соотношение сухих и вегетирующих частей примерно 75/25. Растение находится в старом генеративном возрасте 2–3 года.
Сенильный период. Особи субсенильного возрастного состояния (ss) характеризуются заметным преобладанием процессов отмирания над процессами роста (рис. 7а), примерно в соотношении 80/20. На особи формируется одна розетка с зелеными листьями (до 10–12 шт.), дающая один генеративный побег, развивающий максимально до 2 корзинок. Количество розеток с усохшими листьями может достигать у наиболее стойких растений 13–14 шт., а чаще – это 10–12 розеток. Растение находится в субсенильном возрастном состоянии 2 года.
Сенильные особи (s) состоят из многочисленных розеток с усохшими листьями (рис. 7b), некоторые из них в своей верхней части имеют до 8 зеленых листьев. Растение находится на конечной фазе своей вегетации, поэтому отсутствует смысл в определении соотношения сухих и зеленых частей. Розетки с усохшими листьями плохо структурированы, т.к. у них уже в возрасте g3 начались процессы разрушения усохших листьев и их количество у таких растений подсчитать не представляется возможным. Диаметр особей в этом возрасте может быть до 30–40 см. Растение пребывает в сенильном возрасте не более одного вегетационного периода.

В данных фитоценотических и эдафических условиях, характеризующихся высочайшим дефицитом влаги и максимальной для каждой определенной местности инсоляцией, происходит быстрое протекание онтогенеза, быстрое «старение» (почти нет молодых генеративных растений), интенсивное усыхание листьев, начинающееся у виргинильных растений и продолжающееся в сенильном возрасте и ускоряющее переход в старческие состояния, вплоть до быстрого отмирания, чаще всего, не достигнув настоящего сенильного возраста. Для обеспечения существования на экстремальных местообитаниях у P.echinocephalus обязательно имеет место вегетативное размножение, поэтому существование популяции обеспечивается как семенным, так и вегетативным способами.
Во фриганоидных сообществах Долгоруковской яйлы и Чатырдага, развитых на южных микросклонах, в наиболее засушливые годы (2019 и 2020 гг.) нами были обнаружены растения, которые по морфологическим признакам соответствовали генеративным растениям, но находились в нецветущем состоянии. Этот факт позволяет предположить, что в крайне жестких условиях часть особей популяции переходит в квазисенильное возрастное состояние. Но его возврат для продолжения нормального онтогенеза зависит от экотопических (прежде всего погодных условий): если будут осадки, то нормальный ход онтогенеза продолжится, если нет – произойдет быстрое старение и отмирание, например, в старом генеративном состоянии (g3) или субсенильном возрасте, без прохождения последнего этапа онтогенеза — сенильного возраста.
ЗАКЛЮЧЕНИЕ
- Для Ptilostemon echinocephalus установлен полный цикл онтогенеза, состоящий из 4 периодов (латентный, прегенеративный, генеративный и сенильный) и 11 возрастных состояний, который особи проходят в течение 15–18 лет.
- Основными критериями для дифференциации прегенеративных возрастных состояний являются: количество листьев, морфологическая форма листа, форма края листовой пластинки, количество вегетирующих розеток и число розеток с усохшими листьями; для виргинильного возраста найден дополнительный критерий- процентное соотношение усохших и вегетирующих листьев.
- Основными критериями для дифференциации генеративных возрастных состояний являются: количество генеративных побегов, число корзинок, формирующихся на каждом из них, количество розеток с вегетирующими и усохшими листьями, процентное соотношение сухих и зеленых листьев.
- Дифференциация субсенильного и сенильного возрастных состояний производится по тем же критериям, которые используются для генеративных особей, основной акцент – на соотношение усохших и вегетирующих частей.
- Расшифрована специфика хода онтогенеза листа в ювенильном возрасте и характер реализации онтогенеза в целом, в зависимости от эколого-ценотических условий фриганоидных сообществ и петрофитных степей.
REFERENCES
- Database The Plant List [http://www.theplantlist.org, date of the application: 01.06.2021].
- Database The international compositae alliance – TICA [https://www.compositae.org, date of the application: 01.06.2021].
- Tutin T. G., Heywood V. H., Burges N. A., Moore D. M., Valentine D. H., Walters S. M., Webb D. A. Flora Europaea, 4, 505 p. (Plantaginaceae to Compositae (and Rubiaceae) Cambridge: Cambridge University Press. 1976).
- Red Data Book of the Republic of Crimea. Plants, algae and mushrooms, V. Ena, A. V. Fateryga (ed.). 480 p. (Simferopol: ARIAL, 2015).
- Red Data Book of the Krasnodar Territory. Animals. III edition, Ed. ed. A.S. Zamotailov, Yu. V. Lokhman, B. I. Wolfov, 720 p. (Krasnodar: Adm. Krasnodar. krai, 2017).
- Red Data Book of the Stavropol Territory. Vol. 1: Plants, ed. A. L. Ivanova, 384 p. (Stavropol: Publishing house of JSC «Polygraphservice», 2002).
- Red Book of the Stavropol Territory. Vol. 1: Plants / ed. A.L. Ivanova, 400 p. (Stavropol: Asterisk, 2013).
- Golub V. B., Grechushkina N. A., Sorokin A. N., Nikolaychuk L. F. Plant communities of the class Onosmato polyphyllae-Ptilostemonetea Korzhenevsky 1990 on the Black Sea coast of the Caucasus and the Crimean Peninsula, Vegetation of Russia, 17–18. (2011).
- Ryff L. E. Rare biotopes of erosion-denudation landscapes of southeastern Crimea, Bulletin GNBS, 124 (2017).
- Zadneprovskaya E. Phytocenotic confinement and spatial structure of the Lamyra echinocephala (Willd.) Tamamsch cenopopulation in the Piedmont Crimea, Science Days of the Vernadsky KFU: conf., 7, 349 (Simferopol, 2017).
- Vakhrusheva L. P., Zadneprovskaya E. V. The current state of the Lamyra echinocephala (Willd.) Tamamsch cenopopulation. in phytocenoses of the landscape-recreational park «Bitak» (environs of the city of Simferopol), Problems of steppe studies, 15, 59 (2019).
- Rabotnov T. Issues of studying the composition of cenopopulations for the purposes of phytocenology, Probl. Botany, I, 465 (1950).
- Uranov A., Bogdanova A. G., Grigorieva N. M. Plant cenopopulations (development and relationships), 136 p. (Moscow: Nauka, 1975).
- Smirnova O., Zaugolnova L. B., Ermakova I. M. Plant cenopopulations (basic concepts and structure). 216 p. (Moscow: Nauka, 1976).
- Zlobin Yu. Principles and methods of studying cenotic populations of plants (teaching aid), 147 p. (Publishing house of Kazan University, 198).
- Zlobin Yu., Sklyar V. G., Klimenko A. A. Populations of rare plant species: theoretical foundations and methods of study: monograph, 439 p. (Sumy: University book, 2013).
- GOST 12038-84 Agricultural seeds. Methods for determining germination (with Amendments No. 1, 2). Enter. 1986-07-01, 30 p. (M.: Standartinform, 2011).
