
MORPHOLOGICAL CRITERIA OF ONTOGENETIC STATUS AND POPULATION STRUTURE OF ECBALLIUM ELATERIUM (L.) RICH. ON AZOV BICH OF KERCH PENINSULA (CRIMEA)
МОРФОЛОГИЧЕСКИЕ КРИТЕРИИ ОНТОГЕНЕТИЧЕСКИХ СОСТОЯНИЙ И СТРУКТУРА ПОПУЛЯЦИИ ECBALLIUM ELATERIUM (L.) RICH. НА АЗОВСКОМ ПОБЕРЕЖЬЕ КЕРЧЕНСКОГО ПОЛУОСТРОВА (КРЫМ)
JOURNAL: «SCIENTIFIC NOTES OF V.I. VERNADSKY CRIMEAN FEDERAL UNIVERSITY. Biology. Chemistry» Volume 10 (76), №3, 2024
Publication text (PDF): Download
UDK: [574.3+582.594](477.75)
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Vakhrusheva L. P., T.I. Vyazemsky Karadag Scientific Station – Nature Reserve of the RAS, Kurortnoye, Feodosia, Russian Federation
Kandaurova D. A., A.O. Kovalevsky Institute of Biology of the Southern Seas of RAS, Sevastopol, Russia
TYPE:Article
DOI:https://doi.org/10.29039/2413-1725-2024-10-3-12-31
PAGES:from 12 to 31
STATUS:Published
LANGUAGE:Russian
KEYWORDS: Ecballium elaterium, population, ontogenetic spectrum, spatial structure, ontogenetic structure, vitality, Kerch Peninsula.
ABSTRACT (ENGLISH):
Ecballium elaterium (L.) Rich. is a Mediterranean-Eurasian steppe species with a wide geographic area. In Crimea, the area is disjunctive: the foothills, the southern coast and the Kerch Peninsula. It is listed in the Red Book of the Republic of Crimea [23] with a conservation status of 3. E. elaterium is a well-studied species, but there is no data on the population structure, which is a gap in its study as a protected component of the flora of Crimea. The purpose of this study is to establish the number, spatial structure, clarify the morphological criteria of age states, determine the ontogenetic structure of the new population. high numbers (1924 individuals) found in 2020 on the northern coast of the Azov Sea.
Morphological criteria were found, according to which 4 periods and 8 ontogenetic states were distinguished in ontogenesis. In the pregenerative period plantlet, juvenile and immature plants were divided by the number of leaves, shape and size of leaves. Morphological features of the root system, the degree of development of the above-ground part of the stem and caudex were used from additional features. The generative phase of ontogenesis was distinguished by the presence of generative shoots, individuals g1, g2 and g3 were differentiated by the shape and size of leaves, the number of above-ground shoots, the ratio of green and dry shoots in an individual, additionally — by the nature of changes in the morphology of the root system. In the post-generative phase of ontogenesis, senile plants are characterized by a return of the leaf shape to juvenile plants, a decrease in the size and underdevelopment of leaves, a reduction in the number and length of green shoots (by 6–8 times or more), destructive morphological transformations of the caudex and root system.
According to the ontogenetic spectrum, the population is normal, contains plants of all ages, the age spectrum type is bimodal, with a peak in juvenile and virginile individuals, which means that the population is young. Additional ontogenetic indices allowed us to confirm and expand the ontogenetic characteristics of the population. The indices of restoration and replacement showed average values > 1, which means that self-maintenance in the population is effective. The aging index was the lowest (Ic = 0.19). The age (Δ) and efficiency (ω) indices also showed that the population is young. Thus, in the E. еlaterium population, self-maintenance processes are normal, and it does not depend on diaspores from the outside. In the spatial structure, two levels of aggregation are traced: the 1st shows the mutual arrangement of individuals within the loci of the population and it was determined as random, which is due to the random distribution and germination of seeds. The 2nd one displays the mutual arrangement of loci within the linear population and is typically group and depends on the dynamism of the substrate and the success of fixation of individuals on it. According to the value of the Q criterion (0.395), the population of E. еlaterium is prosperous. The proportion of individuals with a high vitality class is 55 %.
Тhe established duration of ontogenesis and its individual phases, the morphology of the root system and the presence of caudex indicate that life form of Ecballium elaterium is a perennial herbaceous polycarpic plant in natural populations of the Crimean Peninsula.
ВВЕДЕНИЕ
Стратегия мирового сообщества по биоразнообразию определена до 2030 года международным документом Convention on Biological Diversity, в котором важнейшее место отводится комплексу действий по сохранению популяций редких видов растений in situ и отмечается: только в естественных местообитаниях в полной мере проявляются адаптивные популяционные признаки, сформировавшиеся в процессе эволюции [1].
На территории Крымского полуострова, с точки зрения произрастания охраняемых видов, наиболее уязвимыми естественными местообитаниями являются морские побережья. Они в первую очередь уничтожаются при организации пляжей, сооружении глэмпингов, набережных и мест отдыха, а слагающие их геологические породы и природный рельеф этих территорий нередко представляют собой весьма динамичные, в том числе и оползневые структуры, создающие экстремальные условия для произрастающих здесь видов. Именно к таким местам обитания в Крыму преимущественно приурочен объект нашего исследования Ecballium elaterium (L.) Rich. Вследствие уникальной способности растения синтезировать многочисленные биологически активные соединения, у зарубежных ученых он является одним из наиболее излюбленных объектов исследования по поиску новых, фармакологически значимых химических веществ [2–4]. Именно с целью выявления биологически активных тканей и клеток тщательно изучены анатомические особенности Е. elaterium [5, 6, 9] для их дальнейшего биотехнологического использования [7–9]. Из анатомо-морфологических особенностей отметим важность данных об аномалиях в строении и развитии корня [10], а также о специфике морфологии цветка, особенности строения и функционирования которого у E. elaterium оказались необходимы для уточнения общих вопросов эволюции этого органа и приспособлений к опылению [11–14]. Практическую значимость имеют результаты о характере варьирования морфологических признаков вегетативных органов в естественных условиях и при культивировании [15, 16].
Однако, несмотря на разнообразие подходов к изучению Е. elaterium, научные данные о биологии и экологии вида преимущественно ограничиваются очерками в Красных книгах, где полностью отсутствуют сведения о современном состоянии и структуре его природных популяций – крайне необходимые для созологической оценки вида, внесенного в природоохранные документы. Цель настоящего исследования – определить численность, особенности пространственной локализации особей, выявить принадлежность к жизненной форме, морфологические критерии онтогенетических состояний вида и на этой основе определить онтогенетическую структуру популяции Е. elaterium высокой численности, найденной в 2020 г. на северном побережье Азовского моря.
МАТЕРИАЛЫ И МЕТОДЫ
Е. elaterium, как средиземноморско-евразиатский степной вид, имеет широкий географический ареал: Cредиземноморье, Восточная Европа, Малая и Средняя Азия, Кавказ и Африка [17]. В Крыму он дизъюнктивный: предгорье, южный берег и Керченский полуостров. Охраняется в Казантипском и Опукском природных заповедниках, но в целом отмечается, что численность популяций не превышает
50–100 особей [23]. В 2020 году на северных склонах береговых обрывов Керченского полуострова, в 1,8 км к северо-западу от пос. Маяк Глазовского сельского поселения Республики Крым была найдена популяция Е. elaterium высокой численности: 1924 особей формируют линейную популяцию вдоль побережья Азовского моря (Рис. 1).
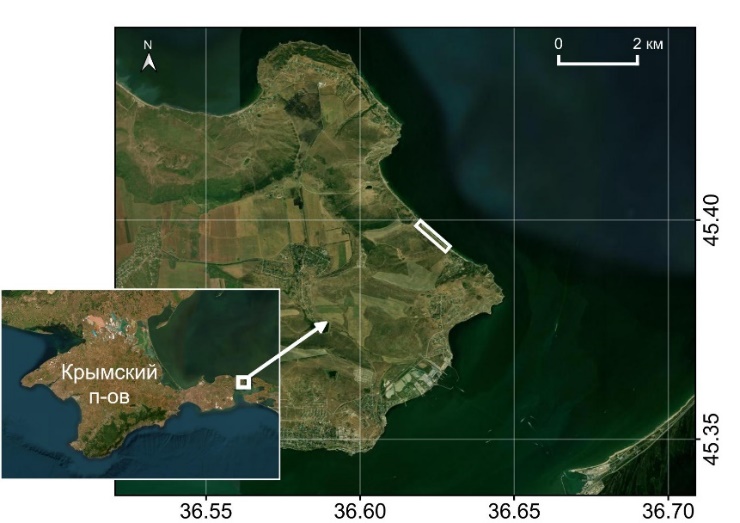
Рис. 1. Локализация популяции Ecballium elaterium на побережье Азовского моря.
Эта территория прибрежной части Азовского моря и Керченского пролива характеризуется сложным балочно-оползневым рельефом, отличающимся высокой динамикой оползневых и осыпных процессов [18]. Е. elaterium преимущественно встречается на береговых абразионных склонах крутизной до 900, сложенных песчанистыми и известняковистыми глинами верхнего миоцена с прослоями карбонатных песчаников [19]. Глины тонко- и средне-слоистые, при выветривании распадающиеся на отдельные пластины, которые образуют осыпи при основании береговых обрывов. Местами они осложнены мелкими оползневыми телами и оплывинами деляпсивного типа. Эти формы рельефа из мелко обломочных и весьма подвижных отложений являются основным субстратом для произрастания популяции Е. elaterium. Скорость смещения обломочных отложений на этом местообитании является лимитирующим фактором, влияющим на возраст отдельных особей, нередко вызывая их катастрофическую гибель (Рис. 2).

Рис. 2. Популяция Е. elaterium на береговых склонах Азовского побережья Керченского полуострова.
Почвенный субстрат с точки зрения химических характеристик имеет следующие особенности: слабощелочной (pH=7,536), средне-гумусный (гумус=3,05 %), повышенной карбонатности (36,05 %), которая блокирует усвоение многих макро- и микроэлементов и усиливает проявление хлора, что может приводить к нарушению баланса потребления почвенных питательных веществ и негативно сказываться на росте растений и снижении для большинства из них возможности произрастания на подобных субстратах (табл. 1). Вероятно, этими особенностями объясняется небольшое число видов, обитающих на береговых склонах данной территории.
Таблица 1
Анализ химического состава грунта в месте произрастания Е. elaterium
| № | EС,
mms |
Na,
мг/л |
Cl, мг/л | K,
мг/л |
Са,
мг/л |
Р, мг/л | Mg, мг/л | N-N03, мг/л |
| 1 | 1,22 | 240,44 | 134,19 | 28,21 | 266 | 3,48 | 42,72 | 1,00 |
Популяция простирается на 820 м как целостное образование, а далее к северо-западу встречаются локусы Е. elaterium, содержащие от 2 до 15 особей, расположенные друг от друга на расстояниях 100–350 м. Популяция представляет практически моноценоз Е. elaterium, поскольку бешеный огурец пружинистый дает здесь до 25 % проективного покрытия из 35 %, характерных для травостоя этой полосы. Единично в составе популяции отмечены еще 8 видов, встречающихся совместно с ним на оползневых склонах: Atriplex patula L., Lactúca serriola L., L. tatarica L., Polygonum robertii Loisel, Сynanchum acutum L., Camelina macrocarpa Wierzb. ex Rchb., Cakile euxina Pobed., Crambe maritima L., количественное участие каждого из них колеблется от единично встречающегося до 1–3 % проективного покрытия. Crambe maritima – вид также внесенный в КК РК – у подножия склона и с выходом на песчаный пляж формирует моноценозные группировки площадью
2–4 м2, которые внедряются в линию популяции Е. elaterium.
По экологическим предпочтениям Е. elaterium отличается широким спектром адаптаций к разнообразным факторам: аэропедофит, гелиофит, мезоксерофит, галофит [22]. Растет группами, что связано со способом распространения семян и особенностями пространственного расположения надземных побегов [23, 24]. В Красную книгу РК [23] Е. elaterium внесен с природоохранным статусом «редкий вид» (3) и характеризуется как «жестко шероховатое растение с простертым или восходящим стеблем», без указания принадлежности к определенной жизненной форме и каких-либо характеристик популяций.
Установление численности, плотности, пространственной структуры, выяснение морфологических критериев возрастных состояний, определение онтогенетической структуры популяции Е. elaterium проводилось стандартными методами [26–28]. Для получения комплексной популяционной оценки определялись индексы восстановления Iв, замещения Iз [29], старения Iс [30], эффективности Iэ [31], возрастности [28], рассчитывалась «дельта-омега» [31] и на их основе устанавливался тип популяции [29].
Пространственная структура изучалась методом «ближайшего соседа», данные анализировались по значению коэффициента неравномерности Кларка-Эванса [32]: – мера того, насколько изучаемое распределение особей отличается от случайного. Дополнительно рассчитывали индекс Одума (IO) , где
D – абсолютная плотность; S – варианса (дисперсия), поскольку IO характеризует распределение особей в каждый момент сбора, поэтому может быть использован в будущем при мониторинговых исследованиях для анализа изменений пространственной структуры популяции [33]. Виталитет популяции определяли по методике Ю. А. Злобина [27]. Учитывая созологическую ценность E. еlaterium и поэтому невозможность измерения фитомассы растений, для оценки виталитетной структуры в качестве детерминирующих признаков были избраны морфологические признаки: длина побегов, их количество, размер листа при основании 3-го от верхушки генеративного побега. Приемом трехмерного ранжирования особи группировались в классы виталитета: а (высший), b (промежуточный), c (низший). В результате устанавливался тип популяции: процветающая (Q=1/2(а+b)>с), депрессивная Q=1/2(а+b)<с) или равновесная (Q=1/2(а+b)=с) [27]. Объем выборки составил 30 особей E. еlaterium генеративного возраста.
![]()
![]()
![]()
![]()
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В изученной популяции были найдены растения всех возрастных состояний и выявлены морфологические признаки, типичные для каждого из них. Е. elaterium в изученной популяции реализует 4 возрастных периода и проходит 8 онтогенетических состояний.
Латентный период, длительность которого у Е. elaterium составляет около одного месяца, растение проводит в виде семян. Семена от коричневых до тёмно-коричневых, эллипсоидные, бочонковидные, гладкие, блестящие, узко-окаймленные, длиной около 3,4 ± 0,2 мм (Рис. 3а). К моменту созревания (середина июля-сентябрь) окружающая их ткань плода превращается в слизистую массу, что приводит к возникновению внутри гидростатического давления до 6 атмосфер, под действием которого плод резко отделяется от плодоножки, а семена, вместе со слизью, с силой выбрасываются на расстояния до 12 м [35].

Рис. 3. Внешний вид Ecballium elaterium: а) семена; б) стадия проростка [Фото авторов].
Прегенеративный период. В первый год жизни бешеный огурец успевает пройти, как правило, две фазы онтогенеза: проростки и ювенильную. Если осень теплая (до конца октября–ноябрь) и длительно задерживаются положительные ночные температуры, отдельные экземпляры могут перейти в имматурную фазу, но такие явления единичны.
Проростки (р) были обнаружены и изучались в естественном фитоценозе.
Прорастание семян подземное, большая часть их появляется спустя месяц после попадания семян в почву (середина августа-сентябрь), но может быть (5–7 % от числа ювенильных особей) и весеннее прорастание (конец апреля-май). Проростки имеют морфологию, типичную для большинства двудольных: семядольные листья с цельной листовой пластинкой, ланцетные, цельнокрайние, верхушка округлая с мало заметной выемкой, сидячие, супротивные до 0,7 ± 0,3 см длины и 0,4 ± 0,3 см ширины (Рис. 3б). Корень стержневой, нитевидный, слабо ветвящийся в нижней трети, длиной до 5,5 см диаметром до 1 мм (Рис. 3б). Вследствие произрастания на постоянно осыпающемся грунте, у проростка хорошо выражен гипокотиль, достигающий в длину до 1,2–2,0 см, но он обнаруживается только при обнажении почвы, т.к. присыпан грунтом.
Переход в ювенильное (j) возрастное состояние происходил примерно через 1 месяц, о чем свидетельствовало появление у проростков двух листьев с листовыми пластинками округло-почковидной формы (0,8 ± 0,4 см длина и 1,0 ± 0,2 см ширина), с неравномерно волнистым краем и короткими черешками: длиной 1,2±0,7 см, диаметром 0,4 ± 0,2 см. К моменту появления листьев ювенильной генерации листья проростков сохранялись зелеными (Рис. 4а). У главного корня в его нижней трети появлялись 2-3 боковых, а главный корень, начиная со средней части, заметно увеличивался в диаметре до 0.6–0.8 см, оставаясь нитевидным на верхушке. В этой же фазе у Е. elaterium развивалась еще одна пара округло-почковидных ювенильных листьев. Большая часть растений (92–98 %) уходили в первую перезимовку в ювенильном возрасте (Рис. 4).

Рис. 4. Ecballium elaterium в ювенильной фазе: а) закладка первой пары ювенильных листьев у проростка; б) зрелое ювенильное растение [Фото авторов].
Подчеркнем также, что у ювенильных особей уже хорошо выражена контрактильность, что обеспечивает к концу вегетации погружение гипокотиля в почву на глубину 3–5 см, и это способствует эффективному закреплению растений на динамичном оползневом грунте к наступлению неблагоприятного времени года (Рис. 5).
Перед уходом в состояние зимнего покоя у ювенильных растений первыми начинают усыхать вторая и третья пары собственно ювенильных листьев, а семядольные долго остаются зелеными и усыхают перед погружением в зимний период покоя.

Рис. 5. Ювенильные растения после смыва грунта штормом [Фото автора].
Имматурные особи (im) появляются весной следующего года. Они имеют яйцевидно-сердцевидные листья (1,3 ± 0,4 см длина; 1,2 ± 0,4 см ширина) со слегка волнистым краем листовой пластинки, заметно выступающей верхушкой и четко выраженными листовыми черешками (1,9 ± 0,6 см длина). У имматурных растений некоторое время сохраняются остатки отмерших за зиму семядольных и ювенильных листьев, но для собственно имматурных особей характерны 4 зелёных листа. Надземная часть стебля еще не выступает над поверхностью почвы, однако хорошо заметна при выкапывании и в этом возрастном состоянии имеет длину до 2,5–3(5) см (Рис. 6).

Рис. 6. Ecballium elaterium в имматурном возрасте [Фото авторов].
Диаметр корня утолщается до 1,0 ± 0,3 см. Утолщение от его нижней трети, типичной для ювенильной фазы, распространяется ближе к корневому основанию, что приводит к заметному оформлению каудекса.
В течение второго вегетационного сезона происходит переход имматурных растений в виргинильное онтогенетическое состояние (v). Виргинильные особи отличаются яйцевидно-сердцевидными слегка лопастными листьями (7, 9 ± 1,2 см длина; 7,7 ± 0,9 см ширина), с городчато-зубчатым краем и уплощенным листовым черешком (0,7 ± 0,3 см ширина). В этот период меняется супротивное листорасположение на очередное. Над поверхностью почвы стебель поднимается на высоту до 1,0–2,5 см, но примерно столько же его длины находится под слоем грунта. Главный корень утолщается, становится мясистым почти по всей длине, достигая в диаметре до 1,5–2,0 см, но по-прежнему остается резко утончающимся к верхушке. У виргинильных особей один боковой корень (редко два), расположенный в средней части главного корня, принимает горизонтальное положение – специфическая морфологическая черта, свойственная большинству растений оползневых экотопов для более надежного закрепления.
По отмеченным признакам виргинильные особи уже вполне соответствуют морфологии взрослого растения (Рис. 7).
 |
 |
Рис. 7. Виргинильное растение Ecballium elaterium в естественном местообитании и в гербарном образце [Фото авторов].
Генеративный период.
В генеративную фазу Е. elaterium обычно переходит только на третий, а чаще на четвертый год жизни. Молодые генеративные особи (g1) отличаются от виргинильных появлением длинных цветоносов (9,1 ± 0,8 см), которые выходят из пазух верхушечных листьев. Листья такой же морфологии, как и у виргинильных растений: жестко-шероховатые, снизу серовойлочные (длина 9,5 ± 0,5 см, ширина 11,8 ± 0,3 см), на мясистых уплощенных черешках (длина 1,3 ± 0,6 см). Стебель поднимается над поверхностью грунта на высоту до 5–10 см, если он выше, то становится лежачим и достигает при этом максимальной длины 36,1 ± 1,4 см. У молодых генеративных особей формируются 1–3 надземных побега, все зеленые. Нижняя часть стеблей у молодых растений всегда погружена в грунт на 3–5 см. Корень уходит на глубину до 25 см, по всей длине мясистый, в самой толстой части достигает 3 см в диаметре, а в верхушечной остается суженным и здесь его толщина не превышает 2 мм. Цветки однополые, однодомные, пентамерные, чашечка спайнолистная, с короткой трубочкой и пятью длинными зубцами; венчик-желтый, спайнолепестный (Рис. 8).
 |
 |
| а | б |
Рис. 8. Внешний вид E. еlaterium в молодой генеративной фазе [Фото авторов].
В зрелом генеративном состоянии (g2) стебель имеет длину до 88, 1 ± 2,7 см, варьируя от 36 до 136 см. В литературе отмечается максимум до 150 см. Благодаря контрактильности, всегда на 12–15 см длиннее, т.к. на эту глубину стебель погружен в почву. Количество живых надземных стеблей 4–9 штук, но уже появляются 1–3 сухих побега. Листья сохраняют яйцевидно-сердцевидную форму с вытянутой верхушкой (длина 11,5 ± 0,6 см, ширина 13,8 ± 0,4 см), но в фазах g2 и g3 край листовых пластинок становится сильнее надрезанным, чем в фазе g1. В зоне, примыкающей к каудексу, корень имеет диаметр 6,0–7,4 см и проникает на глубину до 50 см, а в средней части и нижней трети обычны 2–3 ветвящихся у верхушек боковых корня, имеющих в почве горизонтальное расположение, нередко растущие почти под прямым углом по отношению к главному корню (Рис. 9).
В старом генеративном состоянии (g3) стебель достигал длины 58, 3 ± 4,3 см, количество живых надземных стеблей 4–5 штук, остатков отмерших – до 7 штук, т.е. в этом возрастном состоянии процессы отмирания начинают преобладать над процессами новообразования. Корень достигал диаметра до 10,5 см в зоне, примыкающей к каудексу и проникал примерно на такую же глубину, как и в фазе g2 (50–55 см). В средней части и нижней трети сохраняются 2–3 ветвящихся горизонтальных корня, но диаметр их в этом возрасте увеличивается в 2–3 раза и составляет1,3 – 1,7 см (Рис. 9б).
 |
 |
| а | б |
Рис. 9. E. еlaterium в зрелом (а) и старом (б) генеративном состояниях [Фото авторов].
В постгенеративный период у E. еlaterium достоверно прослеживается только одно онтогенетическое состояние – сенильное. Для сенильных (s) особей характерно развитие 2–4 надземных побегов длиной 5–7 см, одревесневших в погруженной в грунт части, множественные остатки оснований отмерших побегов. Листья округло-почковидной формы, напоминающие листья ювенильных растений (длина 5,3 ± 0,6 см, ширина 6,5 ± 0,5 см), со слегка волнистым краем листовой пластинки, у многих побегов листья недоразвиты (Рис. 10а). Корневая система деформированная, за счет отмерших верхушек корней, а также вследствие одревеснения и частичного отмирания тканей в области основания корня и в зоне каудекса, который уже не имеет способности формировать почки возобновления. Цветоносные побеги не образуются. Растение в этом возрасте пребывает не более
1–2 вегетационных сезонов и погибает (Рис. 10а, б).
 |
 |
| а | б |
Рис. 10. Сенильные особи Е. elaterium [Фото авторов].
На основании выявленных морфологических критериев была проведена дифференциация особей на фрагменте линейной трансекты с подсчетом доли участия каждой онтогенетической группы растений в структуре популяции, что показало: популяция нормальная, полночленная, поскольку содержит особи всех возрастных состояний, тип возрастного спектра бимодальный, с пиком на ювенильных и виргинильных особях, при небольшом преобладании виргинильного пика, т.е. популяция молодая. Такой тип спектра свидетельствует о благополучном ее состоянии и достаточном резерве для будущего существования: особи прегенеративных онтогенетических фаз развития в сумме достигают более 80 % состава популяции (Рис. 11).
Расчет дополнительных онтогенетических индексов: восстановления (Iв), старения (Iс), замещения (Iз), эффективности (ω) и коэффициента возрастности (Δ) позволили подтвердить и расширить онтогенетическую характеристику популяции Е. elaterium (Табл. 2).
Индексы восстановления и замещения показали средние значения (Табл. 2), но каждое из них > 1, значит: на одно генеративное растение приходится более одного потомка и самоподдержание в популяции протекает эффективно [34]. Индекс старения оказался наименьшим (Iс=0,19), что подтвердило данные онтогенетического спектра – число особей постгенеративного периода соответствует 2 % (Рис. 11). Индексы возрастности (Δ) и эффективности (ω) также показали, что популяция является молодой. По принятым современным системам классификации [26, 29], популяция E. еlaterium является нормальной, т.е. процессы самоподдержания протекают в ней нормально, и она не зависит от зачатков извне.
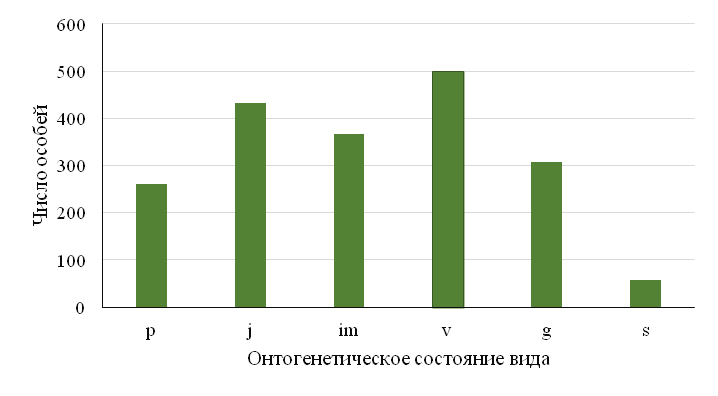
Рис. 11. Онтогенетический спектр Ecballium elaterium.
Таблица 2
Онтогенетические индексы популяции E. еlaterium
| Онтогенетические индексы по Л.А. Жуковой и Н.В. Глотову | Индекс возрастности по А.А. Уранову | Индекс эффективности
по Л.А. Животовскому |
Тип популяции по классификации «дельта-омега» | Тип
популяции по Л.А. Жуковой |
||
| I в | I с | I з | Δ | ω | ||
| 2,6 | 0,19 | 1,3 | 0,13 | 0,43 | Молодая | Нормальная |
Пространственно по всей линии склонов побережья популяция представлена в виде локусов размером 100 – 200 м2 с проективным покрытием E. еlaterium 25 %, которые чередуются с участками такой же площади, где полностью отсутствуют растения или встречаются в количестве 1–7 экземпляров. Пространственная структура внутри локусов, оцененная по значению коэффициентов (коэффициент Кларка Эванса R=4,00; индекс Одума (IO) =1,02), свидетельствует о высокой степени агрегированности особей и групповом их распределении по площади локусов, что, в значительной степени, обусловлено групповым разбрасыванием семян, объединенных слизью плода и, как следствие, групповым прорастанием. Взаимное размещение локусов в пределах линейной популяции является также типично групповым (контагиозным) и определяется преимущественно высокой динамичностью субстрата и успешностью закрепления особей на оползневом грунте. Таким образом, в пространственной структуре популяции имеют место два уровня агрегированности: первый – групповое внутри локусов и второй – контагиозный тип размещения локусов на более стабильных участках склона.
В соответствии с избранными значениями признаков, особи E. еlaterium были распределены по классам виталитета, что показало преобладание растений с высоким классом виталитета: в популяции 55 % особей принадлежат к классу а (Табл. 3).
Таблица 3
Оценка виталитета ценопопуляции E. еlaterium
| Популяция | Доля особей по классам | Тип популяции | ||
| a | b | c | ||
| E. еlaterium | 0,55 | 0,24 | 0,21 | Процветающая |
Значение критерия Q (0,395) подтвердило принадлежность изученной популяции E. еlaterium к процветающему типу.
Дискуссия
В научной литературе, где приводится общее описание признаков Е. elaterium, авторы относят его к летне-зеленым озимым однолетникам [22, 35, 37]. Однако в отечественных и зарубежных публикациях [24, 38–40] есть мнение, что Е.elaterium – это многолетнее травянистое растение. В Красной книге РК [23] в очерке об Е. elaterium деликатно обойден этот нерешенный вопрос, и вид охарактеризован как «жестко шероховатое растение с простертым или восходящим стеблем», без указания принадлежности к определенной жизненной форме. Наши наблюдения в течение 5 лет за длительностью процессов вегетации и особенностями перехода из одного возрастного состояния в другое, а также изучение морфологии подземных побеговых органов (в частности, каудекса) и корневой системы этого растения (Рис. 9, 10) позволяют без сомнения утверждать, что в условиях Крымского полуострова Ecballium elaterium в природных популяциях является многолетним травянистым поликарпиком.
ЗАКЛЮЧЕНИЕ
- Найдена в 2020 г новая популяция Ecballium elaterium высокой численности (1924 особи) на 820 м Азовского побережья.
- В онтогенезе E. еlaterium выделено 4 периода и 8 онтогенетических состояний. Для дифференциации особей всех онтогенетических состояний информативными оказались форма и размер листа. Для разделения на p, j, im, v показаны также число листьев, строение корневой системы, степень развития надземной части стебля и каудекса. Разделение на g1, g2 и g3 проводили по наличию генеративных побегов, количеству надземных побегов, соотношению зеленых и сухих побегов, дополнительно – по характеру изменения морфологии корневой системы. Cенильные растения (s) отличали по деструктивным морфологическим трансформациям всех органов.
- Онтогенетический спектр изученной популяции полночленный, бимодальный; онтогенетические индексы показали, что популяция является молодой, нормальной (самоподдержание осуществляется эффективно, не зависит от зачатков извне), имеет достаточный резерв для будущего существования: особи прегенеративных онтогенетических фаз в сумме составляют более 80 %.
- Пространственная структура контагиозная на протяжении линии популяции и внутри локусов.
- В популяции преобладают растения с высоким классом виталитета а (55 %). Значение критерия Q (0,395) подтверждает принадлежность изученной популяции E. еlaterium к процветающему типу.
- В условиях природных популяций Крыма Е. elaterium по жизненной форме является многолетним травянистым поликарпиком.
Работа выполнена в рамках государственного задания ФИЦ ИнБЮМ по теме «Биоразнообразие как основа устойчивого функционирования морских экосистем, критерии и научные принципы его сохранения» (№ гос. регистрации 124022400148-4).
Работа выполнена в рамках темы НИР «Изучение биотических и абиотических компонентов наземных экосистем, особенности их структурно-временной организации в различных климатических условиях среды» номер темы 124030100098-0 Карадагская научная станция им. Т. И. Вяземского – природный заповедник РАН – филиал Института биологии южных морей им. А. О. Ковалевского РАН
- The Convention on Biological Diversity. – URL: https://www.cbd.int (accessed: 12.06.2024).
- Abbassi F., Ayari B., Mhamdi B. [et al.] Phenolic contents and antimicrobial activity of squirting cucumber (Ecballium elaterium) extracts against food-borne pathogens, Pakistan journal of pharmaceutical sciences, 27 (3), 475 (2014).
- Arslan S., Okur M. H., Zeytun H. [et al.] A new experimental rat model of pancreatitis using Ecballium elaterium, International Journal of Surgery, 23, 160 (2015).
- Karimi N., Bohlooli S., Mazani M. The Evaluation of Cytotoxic Effect of Nanoliposomal Extract of Ecballium Elaterium on Human Gastric Adenocarcinoma (AGS) cell line, Nanomed Res, 1, 9 ( 2016).
- Christodoulakis N. S., Kollia K., Fasseas C. Leaf structure and histochemistry of Ecballium elaterium (L.) A. Rich. (squirting cucumber) Flora-Morphology, Distribution, Functional Ecology of Plants, 206 (3), 191 (2011).
- Rus L., Ielciu I. I., Păltinean R. Morphological and histo-anatomical study of Bryonia alba L.(Cucurbitaceae), Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 43 (1), 47 (2015).
- Attard E. Rapid Detection of Cucurbitacins in Tissues and in vitro Cultures of Ecballium, Cucurbit Genet Coop Rep, 25, 71 (2002).
- Bohlooli S. Jafari N., Jahed S. Cytotoxic effect of freeze-dried extract of Ecballium elaterium fruit on gastric adenocarcinoma (AGS) and esophageal squamous cell carcinoma (KYSE30) cell lines, Gastrointest Cancer, 43, 579 (2012).
- Ielciu I. I. Bryonia alba L. and Ecballium elaterium (L.) A. Rich. – two related species of the cucurbitaceae family with important pharmaceutical potential, Farmacia, 64 (3), С323 (2016).
- Dubravec K. Anomalna sekundarna graa korijena (trcalice (Ecballium elaterium (L.) A. Rich.), Acta Bot. Croat, 26, 151 (1967).
- Fahn A. Shimony A. Nectary structure and ultrastructure of unisexual flowers of Ecballium elaterium (L.) A. Rich. (Cucurbitaceae) and their presumptive pollinators, Annals of Botany, 87 (1), 27 (2001).
- Rust R. W., Vaissière B. E., Westrich P. Pollinator biodiversity and floral resource use in Ecballium elaterium (Cucurbitaceae), a Mediterranean endemic, Apidologie, 34 (1), 29 (2003).
- Gerashchenkov G. A., Rozhnova N. A. Mobile genetic elements in the evolution of sex in plants, Genetics, 46 (11), 1445 (2010).
- Gashkova I. V. Melon collection (Cucumis melo L.): mobilization, study, storage, Problems of studying and preserving the flora of Eurasia, 2, 147 (2017).
- Inan M., Kirici S., Özel A. Investigation of some morphological properties of Echallium elaterium L. in the natural habitat and cultivation conditions, Plants of the Balkan Peninsula: into the next Millennium. The Proceedings of the 2nd Balkan Botanical Congress held at Istanbul, Turkey, 1, 569 (2000).
- This volume contains submitted papers presented at the «Second Balkan Botanical Congress».– URL: https://biore.bio.bg.ac.rs/handle/123456789/2288 (accessed: 24.04.2024).
- PlantNet.– URL: https://identify.plantnet.org/ru/k-worldflora/species/Ecballium%20elaterium%20(L.)%20A.Rich./data (accessed: 27.04.2024).
- Blagovolin N. S. Geomorphology of the Kerch-Taman region,193 р. (Moscow: Academy of Sciences of the USSR, 1962).
- Muratov M. V. Guide to educational geological practice in the Crimea, Moscow: Nedra, 1, 192 (1973).
- Ignatov E. I., Chistov S. V. Ecological and geomorphological assessment of the coast and bottom of the Kerch Strait in connection with solving transport problems, Environmental safety of coastal and shelf zones, 8, 163 (2003).
- Smekalova T. N., Kulikov A. V., Beilin D. V. [et al.] Archaeological-Geophysical sites pamyatnikov in south-eastern parts of Kerch Peninsula, History and archeology of Crimea, 13, 373 (2020).
- Golubev V. N., Biological flora of Crimea, 86 p. (Yalta: NBS-NSC, 1996).
- Red book of the Republic of Crimea. Plants, algae, fungi, Rel. ed. d. b. n., Prof. A. V. Ena and Ph.D. A. V. Vateryga, 480 p. (Simferopol: it «Arial «LLC, 2015).
- The Red Book of the Krasnodar Territory. Plants and mushrooms, Resp. ed. S. A. Litvinskaya. – 3rd ed, 850 p. (Krasnodar, 2017).
- The Red Book of Ukraine. Flora, Ed. Ya. P. Didukh, 900 p. (K.: GlobalConsulting, 2009).
- Rabotnov T. A., Life cycle of perennial herbaceous plants in meadow cenoses, Proceedings of the BIN an USSR. – Ser. 3, Geobotany, 6, 7 (Moscow: USSR Academy of Sciences, 1950).
- Zlobin Yu. A., Sklyar V. G., Klimenko A. A., Populations of rare plant species: theoretical foundations and methods of study: monograph. 439 p. (Sumy: University book, 2013).
- Uranov A. A. Age spectrum of phytocenopopulations as a function of time and energy wave processes, Scientific Reports of the Higher School. Biological Sciences, 2, 7 (1975).
- Zhukova L. A. Dynamics of coenopopulations of field plants in natural communities. In: T. I. Serebriakova (ed.). Dynamics of coenopopulations, 9 (Nauka, Moskow, 1987).
- Glotov N. V. On the assessment of the parameters of the age structure of the plant population, Life of populations in a heterogeneous environment, 1, 146 (1998).
- Zhivotovsky L. A., Ontogenetic States, effective density and classification of plant populations, Ecology, 1, 3 (2001).
- Kharitonov S. P., The «nearest neighbor» Method for mathematical evaluation of the distribution of biological objects on a plane and on a line, Bulletin of the Nizhny Novgorod University. N. I. Lobachevsky. – Ser. Biology, 1, 213 (2005).
- Odum Yu., Ecology, 1, 328 p. (M.: Mir Publishing House, 1986).
- Zhivotovsky L. A., Osmanova G. O. Population biogeography of plants, 128 р. (Yoshkar-Ola: Vertical LLC., 2019).
- Chen Y., Wu Q, Zhang Z. [et al.] Preparation of cucurmin- loaded liposomes and evaluation of their skin permeation and pharmacodynamics, Molecules, 17 (5), 5972 (2012).
- Denisova G. A. The pumpkin family (Cucurbitaceae) (ed. by A.L. Takhtajyan), Plant life, 5 (2), 53 (1981).
- Alabama Plant Atlas, 2021.– URL: https://web.archive.org/web/20220519092034/http://floraofalabama.org/ (accessed:: 24.04.2024).
- Alekhine A. A. Types and varieties of seeds. Cucurbitaceae in the Botanical Garden of Kharkov University, The editorial board, 88, 37 (2001).
- European Regional Development Fund (ERDF) in Malta.– URL: https://pact-for-skills.ec.europa.eu/stakeholders-and-business/funding-opportunities/european-regional-development-fund-erdf-malta_en (accessed: 24.04.2024).
- Keener B. R., Diamond. A. R, Davenport L. J. [et al.]. Alabama Plant Atlas, 274 р. (Alabama : University of West Alabama, 2021).
- Yurtsev B. A. Life forms: one of the nodal objects of botany, Problems of ecological morphology of plants. M.: Nauka, 1, 9 (1976).
- Notov A. A., Zhukova L. A. On the role of the population-ontogenetic approach in the development of modern biology and ecology, Bulletin of Tver State University. Series: Biology and Ecology, 32, 293 (2013).
