
ASSESSMENT OF THE POSSIBILITY OF CORRECTION OF PSYCHOEMOTIONAL DISORDERS IN ALCOHOLIZED RATS WITH STIMULATION OF DOPAMINERGIC TRANSMISSION
ОЦЕНКА ВОЗМОЖНОСТИ КОРРЕКЦИИ ПСИХОЭМОЦИОНАЛЬНЫХ РАССТРОЙСТВ У АЛКОГОЛИЗИРОВАННЫХ КРЫС ПРИ СТИМУЛЯЦИИ ДОФАМИНЕРГИЧЕСКОЙ ПЕРЕДАЧИ
JOURNAL: «SCIENTIFIC NOTES OF V.I. VERNADSKY CRIMEAN FEDERAL UNIVERSITY. Biology. Chemistry» Volume 10 (76), №1, 2024
Publication text (PDF): Download
UDK: 591.5:612.821.4
AUTHOR AND PUBLICATION INFORMATION
AUTHORS:
Balakireva G. A., Donetsk state university, Donetsk, Russia
Balakireva Е. A., Donetsk state university, Donetsk, Russia
TYPE:Article
DOI:https://doi.org/10.29039/2413-1725-2024-10-1-16-34
PAGES:from 16 to 34
STATUS:Published
LANGUAGE:Russian
KEYWORDS: anxiety, depression, motor activity, research activity, emotionality, alcoholization, dopamine, L-DOPA, Sulpiride.
ABSTRACT (ENGLISH):
The purpose of the presented fragment of the complex work is to assess the possibility of compensating for psychoemotional disorders that occur after prolonged ethanol intake in male white rats with drugs that stimulate transmission in the dopaminergic synapse, taking into account the individual characteristics of animal organism.
Method. The experiment is executed on 80 sexually mature rats-males (180–220 g).
The level of anxiety was assessed in an elevated plus maze based on the total time spent by the animal in the open space. Locomotor and exploratory activity, grooming behavior of animals was assessed using open field within 5 minutes. The depressive level of rats was determined in the test of Porsolt by the total time of the time of immobility and the number of periods of immobility for 6 minutes of testing. The number of fecal boluses was tried about emotional animals. After the initial (control) test battery in the above test animals were divided into three subgroups according to the severity of depressive level in the test of Porsolt. Alcoholization was carried out for 14 days by intraperitoneal injection of a solution of ethanol in a 10 % solution at the rate of 2 g/kg of animal weight, after which the animals were again tested. The dopaminergic system was stimulated by introducing a precursor of dopamine synthesis – L-DOPA («Madopar». Roche S.p.A., Italy) at a dose of 50 mg/kg intraperitoneally for 14 days and a dopamine autoreceptor blocker – Sulpiride («Eglonyl», Sanofi Winthrop Industrie, France) at a dose of 10 mg/kg intraperitoneally for 14 days.
Results. In model experiments on male white rats, the possibility of correcting behavioral disorders that occur against the background of two weeks of alcoholism was investigated by introducing an autoreceptor blocker of dopamine (sulpiride) and a precursor to dopamine synthesis (L-DOPA). It was found that both sulpiride and L-DOPA correct the anxiogenic effect of ethanol in animals regardless of their initial anxiety level. L-DOPA makes it possible to neutralize the inhibition of research and motor activity induced by two weeks of ethanol administration in males with initially extreme levels of anxiety, while sulpiride is only in low–anxiety rats. The introduction of a precursor to dopamine synthesis turns out to be more effective for correcting an ethanol-induced increase in the depressiveness index of male white rats.
Conclusion. Thus, can be concluded that the administration of the autoreceptor blocker dafomine sulpiride and the precursor of the synthesis of this mediator L-DOPA to alcoholized males allows to correct the ethanol-induced increase in anxiety in initially medium and highly anxious males, inhibition of research and motor behavior in individuals who showed a low level of anxiety and depressogenic effect of prolonged administration of ethanol in male white rats, regardless of their individual characteristics. features. At the same time, in some cases, L-DOPA implements these effects to a greater extent than sulpiride.
ВВЕДЕНИЕ
Вопрос об эффективности нивелирования расстройств аффективного спектра, возникающих на фоне длительного приема этанолсодержащих продуктов, стоит весьма остро, что обусловлено общими звеньями в механизмах развития как алкоголизма, так и патогенеза психоэмоциональных расстройств [1–5]. Общими звеньями в данном случае выступают ряд нейромедиаторных систем мозга, через которые реализует свои эффекты этанол. В частности – дофаминергическая система головного мозга, нейроны которой, являясь основной мишенью действия психоактивных веществ, изменяют под действием этилового спирта свою активность, где смена усиленной продукции ими дофамина приводит к компенсаторному снижению его содержания в синаптической щели [6–10].
Известно, что в основе патогенеза таких расстройств психоэмоционального спектра как тревожные, двигательные или депрессивные лежит нарушение моноаминовой медиации в головном мозге [11, 12]. Этанол способен модулировать активность катехоламинергических и ГАМК-ергических нейронов, посредством изменения состояния мембраны нейронов, а, следовательно, и их возбудимости, что неизбежно отразится на проявлениях высших функций мозга [13, 14].
В связи со сказанным выше, представляется возможным путем влияния на разные звенья синаптической дофаминергической передачи скомпенсировать некоторые эффекты длительного воздействия этанола на психоэмоциональное состояние животного организма. Вместе с тем, данных, касающихся возможности индивидуального подхода в коррекции этанолиндуцированных расстройств высших функций мозга крайне мало в современной литературе. Решение этого вопроса позволило бы более эффективно оказывать терапевтическую помощь людям.
Таким образом, целью данного фрагмента комплексной работы явилась оценка возможности компенсации психоэмоциональных расстройств, возникающих после длительного приема этанола у самцов белых крыс, с помощью препаратов, стимулирующих передачу в дофаминергическом синапсе, с учетом индивидуальных особенностей животного организма.
МАТЕРИАЛЫ И МЕТОДЫ
Эксперимент выполнен на 80 половозрелых беспородных крысах-самцах массой 180–220г, содержащихся в стандартных клетках в условиях естественного светового режима при свободном доступе к питью и стандартном гранулированном корме. Все исследования были выполнены в соответствии с «Руководством по уходу и использованию лабораторных животных» (публикация Национального института здоровья № 85-23, США) и «Руководством по экспериментальному (доклиническому) изучению новых фармакологических веществ» [15]. Поведенческие эксперименты проводились в первой половине дня. Для оценки таких психоэмоциональных показателей как уровень тревожности, депрессивности и двигательной активности был использован комплекс методов, традиционно применяемый в нейропсихофармакологии.
Уровень тревожности крыс определяли в приподнятом крестообразном лабиринте (ПКЛ) [10] по общему времени пребывания животного на открытом пространстве лабиринта и частоте повторных выходов на него в течение 5 минут тестирования. ПКЛ представляет собой приподнятый над уровнем пола лабиринт, два из четырех рукавов которого по периметру имеют стенки (закрытые рукава), а остальные – нет (открытые рукава). Установка располагается на высоте 80 см над уровнем пола на центральной опоре-ножке и сконструирована из пластика, окрашенного в черный цвет. Линейные размеры рукавов: ширина – 10 см, длина – 45 см, высота стенок в закрытых рукавах – 10 см; в месте пересечения рукавов имеется центральная площадка 10×10 см. При тестировании крысы плавно опускались в центр лабиринта, где визуально регистрировалось их поведение. После каждого животного лабиринт протирался изнутри мокрыми и сухими салфетками, а также дезодорировался раствором этилового спирта.
В норковой камере за 5 минут тестирования оценивали двигательную активность по количеству пересеченных квадратов и исследовательскую активность по суммарному количеству вертикальных стоек и заглядываний в отверстия-норки [16], а так же частоту актов груминга. Норковая камера представляет собой открытый пластиковый ящик с линейными размерами 60×60×40 см, пол которого – квадрат из пластика, выкрашенный изнутри зелено-голубой краской и приподнятый над дном ящика на высоту 3 см. Пол разделен тонкими белыми линиями на 9 равных квадратиков (20×20 см); по периметру квадратиков просверлены отверстия-«норки» диаметром 3 см. При тестировании крыс плавно опускали в центр норковой камеры, где визуально в течение 5 минут регистрировалось их поведение. После каждого животного камера протиралась изнутри мокрыми и сухими салфетками, а также дезодорировалась раствором этилового спирта.
Уровень депрессивности животных устанавливали с помощью теста Порсолта [15]. При тестировании крыс опускали в стеклянный цилиндр емкостью 20 литров и высотой 30 см, наполненный водой (t=27–28 °С). За 6 минут тестирования фиксировали количество и продолжительность периодов полной иммобильности (неподвижности) животного, частоту таких периодов, а также количество фекальных болюсов. Степень выраженности депрессивности определялась с помощью подсчета суммарного времени неподвижности животного, по количеству фекальных болюсов судили об их эмоциональности.
Сульпирид («Eglonyl», Sanofi Winthrop Industrie. France) вводили внутрибрюшинно в течение 14 дней в дозе 10 мг/кг [8], после чего животные снова проходили тестирование. Таким образом, каждое животное было протестировано трижды: в начальных условиях, после хронического введения этанола и после введения сульпирида. L-ДОФА («Madopar», Roche S.p.A., Italy) вводили в течение 14 дней в дозе 50 мг/кг внутрибрюшинно в течение 14 дней [9].
Поведенческое фенотипирование – процедура комплексной оценки поведенческих характеристик лабораторных животных – разработана в целях стандартизации сбора и интерпретации поведенческих данных для выявления индивидуально-типологических отличий начиная с элементарных моторно-двигательных реакций и заканчивая особенностями эмоционально-психической сферы [17]. Предварительное поведенческое фенотипирование представляет собой интерес с той точки зрения, что позволяет установить индивидуальные особенности реагирования лабораторных животных на те или иные воздействия.
Как установлено исследователями [18–20], в основе нейрофизиологических и нейрохимических механизмов, определяющих индивидуально-типологические различия поведения, лежат биохимические особенности организации различных отделов головного мозга. Данный факт позволяет осуществлять более эффективную коррекцию различного рода аффективных расстройств, возникающих на фоне воздействий разного генеза.
После исходного (контрольного) тестирования в батарее вышеуказанных тестов животные были разделены на три подгруппы согласно выраженности тревожности в приподнятом крестообразном лабиринте.
Разделение исследуемой группы животных на подгруппы с различным уровнем тревожности проводилось согласно сигмального отклонения (±0,67δ) [19].
Обработка данных осуществлялась с использованием пакета программ Statistica 6.0. Учитывая, что по результатам предварительной проверки гипотезы о нормальном распределении данных по тесту Колмогорова-Смирнова нормальность не подтвердилась, в дальнейшем в работе использовали непараметрические методы математической статистики (U-критерий Манна-Уитни для независимых переменных). Принятый уровень значимости составлял 0,05.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
В таблице 1 приведены результаты исходного тестирования животных в условиях батареи тестов. Как следует из данных, представленных в таблице, низкотревожные самцы отличаются самыми высокими значениями количества повторных выходов на открытое пространство лабиринта и показателями исследовательской и двигательной активности, частоты актов груминга в норковой камере. Вместе с тем, им свойственна низкая депрессивность и минимальная эмоциональность в тесте Порсолта.
Таблица 1
Показатели поведения у животных в исходных условиях и после длительной алкоголизации (X±m)
| Поведенческие показатели | Этап эксперимента | Уровни тревожности | |||
| высокий (n=26) | средний (n=34) | низкий(n=20) | |||
| Результаты тестирования в приподнятом крестообразном лабиринте | |||||
| Открытые рукава, с | контроль | 0,0# | 42,1±2,36 | 181,5±4,12#* | |
| алкоголизация | 12,9±3,30 | 14,1±3,11● | 0,0● | ||
| Выходы в открытое пространство | контроль | 0,0# | 1,6±0,14 | 2,3±0,15#* | |
| алкоголизация | 0,6±0,12 | 0,8±0,18● | 0,0● | ||
| Результаты тестирования в открытом поле | |||||
| Исследовательская активность | контроль | 11,1±0,94 | 11,8±0,89 | 15,0±1,05#* | |
| алкоголизация | 2,4±0,93● | 6,8±0,73● | 2,5±0,68● | ||
| Двигательная активность | контроль | 14,9±1,15 | 16,1±1,19 | 18,0±1,28#* | |
| алкоголизация | 5,4±0,85● | 16,0±1,17 | 7,5±0,86● | ||
| Частота актов груминга | контроль | 0,9±0,21 | 1,1±0,33 | 3,0±0,21#* | |
| алкоголизация | 0,9±0,46 | 0,7±0,37 | 1,3±0,25● | ||
| Результаты тестирования в тесте Порсолта | |||||
| Время неподвижности, с | контроль | 28,0±2,88 | 27,1±1,75 | 18,5±2,03#* | |
| алкоголизация | 76,3±5,26● | 79,2±3,09● | 86,8±6,14● | ||
| Общее количество периодов неподвижности | контроль | 5,9±0,75 | 5,6±0,57 | 4,8±0,61 | |
| алкоголизация | 12,3±0,73● | 13,3±0,65● | 15,3±0,91● | ||
| Количество фекальных болюсов | контроль | 3,0±0,43 | 3,3±0,36 | 1,3±0,25#* | |
| алкоголизация | 9,1±0,51● | 8,6±0,41● | 6,8±0,38● | ||
Примечание: # – различия статистически значимы в сравнении показателей условного контроля (средний тип выраженности показателей поведения) с группами высокого и низкого типа показателей поведения; * – различия статистически значимы при сравнении показателей группы с крайними типами выраженности показателей поведения; ● – различия статистически значимы при сравнении значений показателей, полученных после алкоголизации с исходными (контрольными).
Самцы, показавшие высокий и средний уровень тревожности в контроле, не отличаются по исследовательской, двигательной и груминговой активности, а так же показателям депрессивности и эмоциональности.
Кроме того, у крыс всех подгрупп тревожности частота актов неподвижности в тесте Порсолта была одинаковой.
Двухнедельная алкоголизация выявила следующие эффекты на психоэмоциональное состояние самцов, отличающихся в контроле уровнем тревожности. Установлено, что высокотревожные крысы не выявили чувствительности в приподнятом крестообразном лабиринте к алкоголизации: суммарное время пребывания на открытом пространстве и количество повторных выходов на него у самцов данной подгруппы достоверных изменений не претерпели (см. табл. 1). У животных с исходно средним уровнем тревожности численные значения маркерного показателя в данном тесте (время, которое крысы проводили на открытых рукавах и центральной платформе) сократилось в 3 раза (p<0,05), а частота повторных выходов на открытое пространство уменьшилась в 2 раза (p<0,05). У низкотревожных в контроле самцов не выявлено повторных выходов на открытое пространство ПКЛ после алкоголизации, а время пребывания на нем сократилось до 0 (p<0,05).
Таким образом, длительная алкоголизация приводит к анксиогенному эффекту у крыс с исходно средним и низким уровнем тревожности.
В норковой камере у всех животных, не зависимо от их исходного уровня тревожности, наблюдалось угнетение исследовательской активности на фоне алкоголизации. При этом, у среднетревожных самцов суммарное количество вертикальных стоек и заглядываний в отверстия-норки после введения этанола сократилось в 1,7 (p<0,05) раза, в то время как у крыс с крайними уровнями тревожности (высоким и низким), количество поведенческих актов, отражающих исследовательскую активность, сократилось в 4,6 (p<0,05) и 6 (p<0,05) раз соответственно. Двигательная активность среднетревожных самцов не изменилась после двухнедельной алкоголизации, а у крыс с исходно низким и высоким уровнем тревожности – сократилась в 2,4–2,8 (p<0,05) раза.
Хроническая алкоголизация привела к сокращению груминговой активности в 2,3 (p<0,05) раза у нетревожных самцов, в то время как у средне- и высокотревожных животных этанол не вызвал изменений в частоте актов груминга.
Обращает на себя внимание тот факт, что направленность изменений показателей поведения, которые отражают депрессивность животных, не зависит от уровня тревожности крыс: у самцов всех исходных подгрупп на фоне двухнедельной алкоголизации установлено увеличение суммарного времени неподвижности в 2,7–4,7 (p<0,05) раза и количества актов замирания в 2,1–3,2 (p<0,05) раза в тесте Порсолта, что указывает на рост депрессивности у них.
Эмоциональность, определяемая по количеству фекальных болюсов, после алкоголизации увеличивалась в 2,6–5,2 (p<0,05) раза у крыс независимо от их исходного уровня тревожности.
Анализ влияния стимулирования дофаминергической системы путем введения алкоголизированным самцам предшественника синтеза дофамина L-ДОФА и блокирования ауторецепторов дофамина сульпирида показал следующие результаты.
Прежде всего, следует отметить, что увеличение количества дофамина в синаптической щели с помощью фармакологических агентов различного механизма действия привело к снижению тревожности хронически алкоголизированных самцов, не зависимо от исходного уровня их тревожности. При этом, анксиолитический эффект в большей степени был выражен при введении L-ДОФА: время пребывания на открытом пространстве лабиринта у высоко- и среднетревожных крыс значительно превысило (более, чем в 5 раз, p<0,05) численные значения данного показателя в исходных условиях (рис. 1, Б), а у низкотревожных животных достигло контрольных значений.
Блокирование ауторецепторов дофамина сульпиридом привело к увеличению (p<0,05) времени нахождения на открытом пространстве лабиринта до исходных значений у среднетревожных самцов (см. рис. 1, А). Нетревожные алкоголизированные самцы показали сокращение тревожности (в 2,4 раза, p<0,05) после введения сульпирида (p<0,05), однако исходных значений оно не достигло.
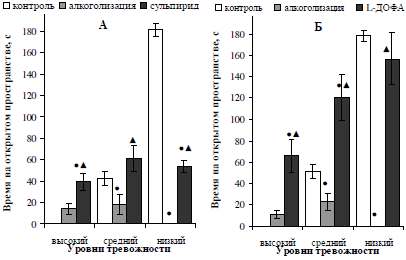
Рис. 1. Характер влияния двухнедельной алкоголизации и последующего введения сульпирида (А) и L-ДОФА (Б) на время пребывания на открытом пространстве лабиринта самцов белых крыс, отличающихся уровнем тревожности.
Примечание: ● – отличия статистически достоверны при сравнении с исходными значениями; ▲ – отличия статистически достоверны при сравнении показателей алкоголизации и введения фармакологических препаратов
Другой показатель тревожности – количество повторных выходов на открытое пространство – у низкотревожных самцов под воздействием используемых фармакологических препаратов изменялся однонаправлено (рис. 1, А, Б): у животных данной подгруппы наблюдалось восстановление частоты выходов до исходных значений (p<0,05). Численные значения данного показателя у среднетревожных алкоголизированных самцов на фоне инъекций сульпирида возросли (p<0,05) и превысили контрольные значения показателя (см. рис. 2, А), что подтверждает анксиолитический эффект блокирования ауторецепторов дофамина у крыс данной подгруппы. Вместе с тем, количество повторных выходов на открытое пространство ПКЛ у алкоголизированных животных с исходно высоким и средним уровнем тревожности после введения предшественника синтеза дофамина не изменилось относительно значений, полученных после двухнедельного введения этанола.
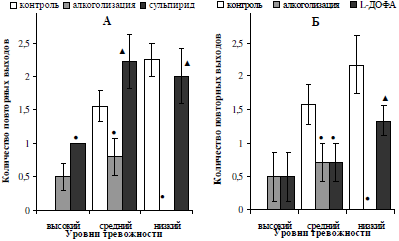
Рис. 2. Характер влияния двухнедельной алкоголизации и последующего введения сульпирида (А) и L-ДОФА (Б) на количество повторных выходов на открытое пространство лабиринта самцов белых крыс, отличающихся уровнем тревожности.
Примечание: ● – отличия статистически достоверны при сравнении с исходными значениями; ▲ – отличия статистически достоверны при сравнении показателей алкоголизации и введения фармакологических препаратов.
Анализ изменения показателей поведенческой активности алкоголизированных самцов, отличающихся по уровню тревожности, в ответ на стимуляцию дофаминергической системы введением предшественника синтеза дофамина L-ДОФА и блокатора ауторецепторов сульпирида, выявил следующее. Прежде всего, установлено, что как сульпирид, так и L-ДОФА скомпенсировали (p<0,05) угнетение исследовательской (рис. 3) и двигательной (рис. 4) активности у алкоголизированных особей, показавших низкий уровень тревожности в исходном тестировании. На данный факт указывает увеличение численных значений исследовательской активности у низкотревожных крыс после инъекций сульпирида в 4,1 раза (p<0,05) и L-ДОФА в 3,2 раза (p<0,05), а так же увеличение количества пересеченных квадратов в 2 (p<0,05) и 1,6 (p<0,05) раза на фоне введения блокатора ауторецепторов дофамина и предшественника его синтеза, соответственно. При этом, двигательная активность нетревожных алкоголизированных крыс после введения сульпирида восстановилась до исходных значений (см. рис. 4, А).
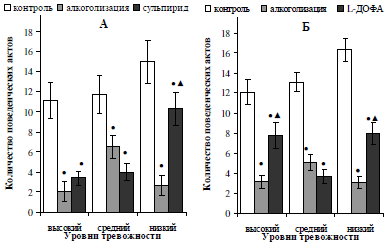
Рис. 3. Характер влияния двухнедельной алкоголизации и последующего введения сульпирида (А) и L-ДОФА (Б) на исследовательскую активность самцов белых крыс, отличающихся уровнем тревожности.
Примечание: ● – отличия статистически достоверны при сравнении с исходными значениями; ▲ – отличия статистически достоверны при сравнении показателей алкоголизации и введения фармакологических препаратов
Обращает на себя внимание тот факт, что использование предшественника синтеза дофамина L-ДОФА оказалось эффективным для восстановления показателей поведенческой активности алкоголизированных самцов с исходно высокой тревожностью, чем применение сульпирида. Так, у животных данной подгруппы установлено увеличение исследовательской активности в 3,3 (p<0,05) раза и двигательной – в 3,6 (p<0,05) раза относительно показателей, полученных после введения этанола (см. рис. 3, 4, Б). В случае двигательной активности наблюдалось ее полное восстановление до контрольных значений.
Среднетревожные алкоголизированные самцы в основном не проявили чувствительности к действию дофаминомиметиков: показатели их поведенческой активности оставались на уровне тех, что были получены после введения этанола. Исключение составило сокращение проявлений двигательной активности в 2,1 (p<0,05) раза у особей со средним уровнем тревожности после введения сульпирида.
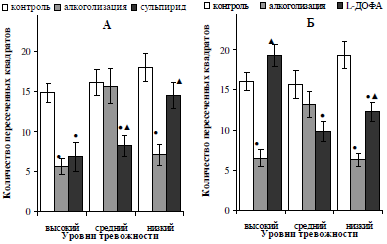
Рис. 4. Характер влияния двухнедельной алкоголизации и последующего введения сульпирида (А) и L-ДОФА (Б) на двигательную активность самцов белых крыс, отличающихся уровнем тревожности.
Примечание: ● – отличия статистически достоверны при сравнении с исходными значениями; ▲ – отличия статистически достоверны при сравнении показателей алкоголизации и введения фармакологических препаратов.
Характер изменения груминговой активности алкоголизированных самцов, отличающихся по исходному уровню тревожности, под действием дофаминомиметиков разного механизма действия в большинстве случаев не дал возможности для установления каких-либо закономерностей вследствие большой дисперсии полученных результатов. Выявлено некоторое стимулирующее действие L-ДОФА (в 4,4 раза (p<0,05)) на груминговое поведение высокотревожных самцов и восстановление груминговой активности у нетревожных крыс после введения сульпирида (p<0,05).
Изменение депрессивно-подобного поведения алкоголизированных самцов под влиянием сульпирида и L-ДОФА выявило однонаправленные изменения показателей поведения в тесте Порсолта, проявившиеся в сокращении показателя депрессивности у крыс всех подгрупп тревожности (рис. 5). При этом, блокирование ауторецепторов дофамина привело к полному восстановлению исходных значений депрессивности (p<0,05) в тесте Порсолта у всех алкоголизированных крыс (см. рис. 5, А), в то время как антидепрессивное действие предшественника синтеза дофамина зависело от исходного уровня тревожности лабораторных животных: чем более низким был исходный уровень тревожности особей, тем в большей степени под действием L-ДОФА у них сокращалось значение депрессивности (см. рис. 5, Б). Так, у высокотревожных алкоголизированных животных суммарное время неподвижности после введения L-ДОФА сократилось в 1,7 (p<0,05) раза и достигло исходных значений, у средне- и низкотревожных – в 2,9 (p<0,05) и 9,7 (p<0,05) раза, соответственно, что в 1,4–2,1 (p<0,05) раза ниже исходных значений данного показаетля.
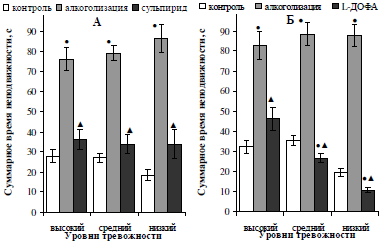
Рис. 5. Характер влияния двухнедельной алкоголизации и последующего введения сульпирида (А) и L-ДОФА (Б) на суммарное время неподвижности самцов белых крыс, отличающихся уровнем тревожности.
Примечание: ● – отличия статистически достоверны при сравнении с исходными значениями; ▲ – отличия статистически достоверны при сравнении показателей алкоголизации и введения фармакологических препаратов.
Динамика изменения частоты замираний в тесте Порсолта у исследуемых подгрупп алкоголизированных крыс после введения сульпирида полностью совпадала с изменением общего времени неподвижности: блокирование рецепторов обратного захвата привело к сокращению показателя (p<0,05) до исходных значений (рис. 6, А). Изменение количества актов неподвижности у алкоголизированных самцов после введения им предшественника синтеза дофамина наблюдалось только в подгруппе низкотревожных особей: частота их замираний сократилась в 1,8 (p<0,05) раза и достигла контрольных значений (см. рис. 6, Б).
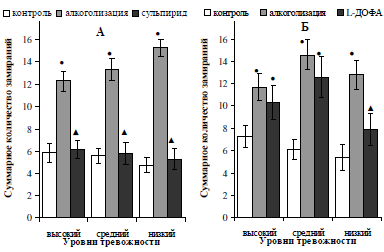
Рис. 6. Характер влияния двухнедельной алкоголизации и последующего введения сульпирида (А) и L-ДОФА (Б) на количество актов неподвижности самцов белых крыс, отличающихся уровнем тревожности.
Примечание: ● – отличия статистически достоверны при сравнении с исходными значениями; ▲ – отличия статистически достоверны при сравнении показателей алкоголизации и введения фармакологических препаратов
Характер изменения эмоциональности у алкоголизированных разнотревожных самцов после введения обоих видов стимуляторов активности дофаминергической системы показал практически одинаковые результаты. Так, низкотревожные алкоголизированные крысы не проявили чувствительности по данному показателю к введению сульпирида и L-ДОФА (рис. 7). Эмоциональность высоко- и среднетревожных особей на фоне введения фармакологических агентов сокращалась в разной степени: у тех животных, которые показали в контрольном тестировании высокую тревожность, количество фекальных болюсов сократилось (p<0,05) до исходных значений, а у тех, которые выявили средние значения тревожности, сульпирид восстановил (p<0,05) исходные значения эмоциональности (см. рис 7, А); инъекции L-ДОФА (см. рис 7, Б) привели к угнетению данного показателя ниже исходных значений в 5,4 (p<0,05) раза.
Усиление тревожности, полученное нами у самцов с исходно средним и низким уровнями тревожности (см. рис. 1, 2), может быть обусловлено резким падением дофамина в прилежащем ядре [21] и эффектами этанола на систему ГАМК-ергических нейронов, на что указывают имеющиеся в литературе данные [22].
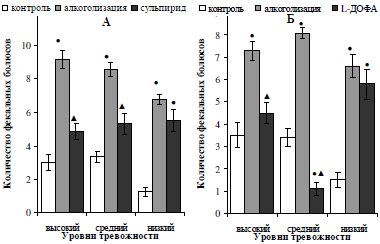
Рис. 7. Характер влияния двухнедельной алкоголизации и последующего введения сульпирида (А) и L-ДОФА (Б) на эмоциональность самцов белых крыс, отличающихся уровнем тревожности.
Примечание: ● – отличия статистически достоверны при сравнении с исходными значениями; ▲ – отличия статистически достоверны при сравнении показателей алкоголизации и введения фармакологических препаратов.
Значительное угнетение исследовательской (в 1,7–6,0 раза (p<0,05)) и двигательной (2,1–2,8 раза (p<0,05)) активности, полученные нами у самцов белых крыс после длительной алкоголизации в норковой камере (см. рис. 3), подтверждают результаты исследований других авторов [3, 23–26]. Такие эффекты могут являться результатом этанолиндуцированного нарушения синаптической пластичности, изменения структуры дендритного дерева, а так же активизации процессов нейродегенерации и угнетения содержания нейротрофического фактора и фактора роста нервов в гиппокампе и префронтальной коре, на что указывают результаты исследований ряда авторов [27, 28]. Эти же являения могут лежать и в основе увеличения показателя депрессивности в тесте Порсолта (см. рис. 5) под воздействием этанола [28, 29]. Вместе с тем, исследования Башкатова В. Г. [30] и Кудрина В. С. [31] указывают на возможность увеличения численных значений двигательного и исследовательского поведения у лабораторных грызунов при введении этанола, что противоречит полученным нами и другими лабораториями, результатам.
Индуцированное длительным введением этанола увеличение эмоциональной нестабильности (см. рис. 7), полученное в наших исследованиях, подтверждает имеющиеся в литературе данные [3]. Однако, некоторые исследования указывают на отсутствие связи между алкоголизацией и эмоциональностью лабораторных грызунов [24].
Влияние препаратов, стимулирующих дофаминергическую передачу путем или блокировнаия рецепторов обратного захвата, или увеличения содержания предшественника синтеза дофамина, оказывается эффективным в ряде случаев при коррекции этанолиндуцированных расстройств психоэмоционального спектра по причине увеличения количества данного медиатора, что восполняет недостаточность дофаминергической системы, возникшей в ответ на хроническую алкоголизацию. На подобный эффект блокаторов ауторецепторов к дофамину после влияния этанола в своих исследованиях указывали некоторые исследователи [21]. Вместе с тем, в литературе имеются данные, что этанол может быть причиной гибели какой-то части дофаминергических нейронов [32] и, соответственно, компенсация их функции способствует нивелированию ряда нарушений аффективной сферы, вызванных этанолом.
В своих предыдущих работах мы уже указывали на тот факт, что длительная алкоголизация значительно увеличивает депрессивность и эмоциональность лабораторных животных независимо от их исходного уровня активности или депрессивности в открытом поле, что может компенсироваться последующим введением препарата, усиливающего дофаминергическую передачу [33, 34].
Подводя итог сказанному, можно сделать вывод, что введение блокатора ауторецепторов дофамина сульпирида и предшественника синтеза данного медиатора L-ДОФА алкоголизированным самцам позволяет скорректировать этанолиндуцированное увеличение тревожности у исходно средне- и высокотревожных самцов, угнетение исследовательского и двигательного поведения у особей, показавших низкий уровень тревожности и депрессогенный эффект длительного введения этанола у самцов белых крыс независимо от их индивидуальных особенностей. При этом, в ряде случаев L-ДОФА в большей степени, чем сульпирид, реализует данные эффекты.
ЗАКЛЮЧЕНИЕ
- Длительная алкоголизация оказала анксиогенный эффект на самцов с исходно средним и низким уровнем тревожности, на что указывает сокращение времени пребывания на открытом пространстве приподнятого крестообразного лабиринта у животных более, чем в 3 раза (p<0,05). Увеличение тревожности алкогоизированными особями данных подгрупп частично компенсировалось введением сульпирида и полностью – введением L-ДОФА. Предшественник синтеза дофамина позволил снизить индуцированную этанолом тревожность в значительно большей степени (более, чем в 5 раз (p<0,05)) относительно значений, полученных после алкоголизации.
- Четырнадцатидневная алкологизация привела к угнетению проявлений исследовательской и двигательной активности более, чем в 2 раза (p<0,05) в норковой камере у всех подопытных животных независимо от их исходного уровня тревожности, показанного ими в ПКЛ. Последующее введение сульпирида позволило несколько (в 2,1–3,7 раза (p<0,05)) скомпенсировать данный эффект у крыс с исходно низким уровнем тревожности, а введение
L-ДОФА повысило численные значения данных показателей у исходно высоко- (в 1,8–3,5 раза (p<0,05)) и низкотревожных (в 1,6–3,2 раза (p<0,05)) особей. - Введение этанола в течение 14 дней увеличивает суммарное время неподвижности в тесте Порсолта у всех экспериментальных животных, независимо от их исходного уровня тревожности в 2,7–4,1 раза (p<0,05). Последующее введение блокатора ауторецепторов дофамина сульпирида полностью восстанавливает исходные значения данного показателя. Введения предшественника синтеза дофамина алкоголизированным самцам возвращает к исходному уровню показатель депрессивности у высокотревожных крыс (p<0,05), и снижает (более, чем в 3,5 раза p<0,05) показатель депрессивности у средне- и низкотревожных самцов, что делает их менее депрессивными, чем при исходном тестировании.
- Длительная алкоголизация увеличивает проявления эмоциональности у животных, независимо от их исходного уровня тревожности, более чем в 2,5 раза (p<0,05). Последующее введения сульпирида нивелирует данный эффект этанола на подгруппу самцов с исходно высоким и средним уровнем тревожности. Введения L-ДОФА нивелирует этанолиндуцированную стимуляцию эмоциональности у высокотревожных самцов и значительно сокращает ее у среднетревожных животных (в 5,4 раза (p<0,05)). На индуцированное этанолом увеличение эмоциональности низкотревожных особей оба препарата эффекта не оказали.
- Pursanov K. A., Homutov A. E., Slobodenuk V. S., Bochkareva A. V. Influence of heparin on rats’ hypodynamia caused by ethyl alcohol. Medicinskij a’manah, 1 (6), 127 (2009) (In Russian)
- Spasov A. A., Petrov V. I. et al. Pharmacological activity of a complex magnesium-containing preparation based on mineral bishofit and pyridoxine hydrochloride studied on the model of chronic alcohol intoxication in rats. Russian Journal of Experimental and Clinical Pharmacology, 66 (5), 40 (2003) (In Russian)
- Pahomova A. O., Kovalenko O. A., Govoruha T. M., Baban V. M., Makarchuk M. Yu. Change of behavioral reactions and lipopersdation processes in liver in strongly alcoholised rats under introduction of quercetin during 14 days, Physics of the alive, 16 (1), 105 (2008) (In Ukrainian).
- Tyurenkov I. N., Voronkov A. V., Borodkina L. E. Effect of phenibut on the behavior of experimental animals under conditions of voluntary chronic alcoholism. Russian Journal of Experimental and Clinical Pharmacology, 68 (3), 42 (2005) (In Russian).
- Bashkatova V. G., Sydakov S. K., Trigub M. M., Bogdanova N. G., Kolpakov A. Introduction of an agonist the kappa-opioid receptors prevents changes of a physical activity and the metabolism of rats caused by acute administration of ethanol. Akademicheskiy zhyrnal Zapadnoy Sibiri, 1 (50), 57 (2014). (In Russian)
- Pivovarchik M. V. Participation of opioid and dopamine systems of the brain in the implementation of the addictive properties of ethanol. Zhurnal GGMU, 4, 3 (2003) (In Russian).
- Lebedev A. A., Droblenkov A. V, Shabanov P. D. Cell Reaction of the Brain Mesocorticolimbic Dopaminergic System on Chronic Alcoholization in Rats, Psychopharmacol biol narcol, 8 (3-4). 2453 (2008). (In Russian)
- Shabanov P. D., Lebedev A. A., Mesherov Sh. K., Pavlenko V. P. Dopamine role in formation of emotional behavior. Obzori po clinicheskoy farmacologii i eksperimentalnoi terapii, 2 (1), 23 (2003) (In Russian)
- Bogdanova I. V. The role of dophamine in mechanisms of forming some disorders of central nervous system and dependent states (Review) Ukrainskiy visnik psyhonevrologii, 19 (67), 5 (2011) (In Russian)
- Kudryavtseva N. N., Bakshtanovskaya I. V., Avgustinovich D. F., Koryakina L. A., Serova L. I., Wishnivetskaya G. B., Kaledin V. I., Popova N. A., Ilnitskaya S. I., Kolesnikova L. A., Amstislavskaya T. G., Gorbach O. V., Lipina T. V., Shigantsov S. N., Madorskaya I. A., Denisova T. F. Social defeats, depression and anxiety: an experimental model. (Novosibirsk: Institute of Cytology and Genetics SD RAS, 1995).
- Shabanov P. D. Psihofarmakologija (Psychopharmacology), 416p. (St. Petersburg: Elbi- StP, 2008). (In Russian)
- Krasnov V. N. Problems of modern diagnostics of a depression. Neuroscience and Behavioral Physiology, 112 (11), 3 (2012) (In Russian)
- Tabakoff B., Hofman L. Neurobiological effects of alcohol. Voprosi narcologii, 5, 27 (2003) (In Russian)
- Shabanov P. D. Narkologiya, 832p. (M.: GEOTAR-Media, 2015) (In Russian).
- Fisenko V. P. Rukovodstvo po eksperementalnomu (doklinicheskomu) izucheniyu novih farmakologicheskich veshestv, 398 р. (Moscow: Minzdrav RF, ZAO «IIA «Remedium»»; 2000). (In Russian)
- Buresh Ja., Bureshova O., Huston D. P. Metodiki i osnovnye jeksperimenty po izucheniju mozga i povedenija (Techniques and the basic experiments for the study of a brain and behavior), 399p. (Moskow: Higher School, 1991). (In Russian)
- Amikishieva A. V. Behavioral phenotyping: up-to-date methods and equipment, Proceedings VOGiS, 13 (3), 259 (2009). (In Russian).
- Sergutina A. V. The effects of L-DOPA on glutamate dehydrogenase activity in the cerebral neurons of rats with different motor activities, Neurochemical Journal, 4 (1), 25 (2010).
- Shalyapina V. G., Vershinina E. A., Rakitskaya V. V., Rizhova L. Yu. Alteration of Active and Passive Wistar Rats Adaptive Behavior in Water-Immersion Model of Depression, I. P. Pavlov Journal of Higher Nervous Activity, 36 (4), 543 (2006). (In Russian).
- Sapronov N. S., Fedotova Yu. O. Effect of L-tryptophan on active avoidance response in male rats with increased testosterone level. Bulletin of Experimental Biology and Medicine, 7, 67 (2000) (In Russian).
- Droblenkov A. V., Shabanov P. D. Neuroglial interactions in dopaminergic structures of the brain in subjects dead from alcohol intoxication. Narcology, 13, 43 (2011) (In Russian).
- Kucher E. O., Egorov A. Y., Filatova E. V., Kulagina K. O. Combine caffeine and ethanol consumption increases alcohol preference in female rats. Narcology, 12, 37 (2010) (In Russian).
- Oskolok L. N., Terentiev A. A. Pathophysiological aspects of chronic alcoholism, drug addiction and toxicomania, Fundamental research, 10, 340 (2011). (In Russian).
- Goldina I. A., Markova E. V., Goldin B. G., Knyazheva M. A., Gaidul K. V. The turmeric protective properties at ethanol-induced behavioral disorders, Saratov Journal of Medical Scientific Research, 13 (1), 131 (2017). (In Russian).
- Kurkin D. V., Morkovin E. I., Osadchenko N. A., Knishova L. P., Bakulin D. A.,. Abrosimova E. E., Gorbynova Yu. V., Turenkov I. N. Correction of psychological and neurological signs of alcohol hangover in rats with acetylcysteine, Pharmacy & Pharmacology, 7 (5), 291 (2019).
- Khomutov A. E., Lizunova A. S. Effect of heparin and ethanol on orientation exploratory behavior of rats in the Open field test, Journal of Siberian Medical Sciences, 2, 42 (2020). (In Russian).
- Fernandez G. M., Stewart W. N., Savage L. M. Chronic drinking during adolescence predisposes the adult rat for continued heavy drinking neurotrophin and behavioral adaptation after long-term, continous ethanol exposure, PLoS One, 11 (3), 1 (2016).
- Bazovkina D. V., Kondaurova E. M., Tsybko A. S., Ilchibaeva T. V., Naumenko V. S., Kovetskaya A. I. The effects of chronic alcoholization on the expression of brain-derived neurotrophic factor and its receptors in the brains of mice genetically predisposed to depressive-like behavior, Molecular Biology, 51 (4), 571 (2017).
- Galecki P., Talarowska M., Anderson G., Berk M., Maes M. Mechanisms underlying neurocognitive dysfunctions in recurrent major depression, Med. Sci. Monit, 21, 1535 (2015).
- Bashkatova V. G., Bogdanova N. G., Nazarova G. A., Alexeeva E. V., Sudakov S. K. The study of the repeated introduction of ethanol in various concentrations on the level of motor activity in rats, Academic Journal of West Siberia, 14, 24 (2018). (In Russian)
- Kudrin V. S., Nadorova A. V., Narkevich V. B., Kolik L. G. An analysis of the behavioral and neurochemical effects of himantane on the dynamics of the ethanol-induced hyperlocomotor response in DBA/2 mice, Neurochemical Journal, 35, 62 (2018). (In Russian)
- Droblenkov A. V., Bobkov P. S., Fedorov A. V., Magradze R. N. Reactive changes of orexynergic and dopamineragic neurons when forming alcohol dependence. Medicina I obrazovanie, 2 (2), 43 (2018) (In Russian)
- Frolova G. A. Behavioural effects of blocking of D2/D3-receptors of dopamine at alkogolization males of white rats with the different level of behavioural activity. Krimskii zhyrnal eksperimentalnoi i klinicheskoi medicini, 7 (4). 75 (2017) (In Russian).
- Frolova G. A. The sulpiride correction of behavioral disorders in alcoholized white male rats with different degrees of depression, Pathological physiology and experimental therapy, 2, 19 (2019). (In Russian).
